THE TRILOBITA
THE trilobites (fig. 1) are arthropods of particular interest because of their great antiquity. They appear on the geological scene at the very beginning of the Paleozoic already fully developed as trilobites; they flourished during the Cambrian and Silurian periods, and continued in diminishing numbers through the Carboniferous into the Permian. Several thousand species have been described, referred to numerous genera and many families in four recognized orders. Such highly organized and diversified animals, therefore, must have had a long evolutionary history in Pre-Cambrian times, though the rocks of this period have so far furnished no evidence of their existence. The trilobites may truly be said to be the oldest of known arthropods, and in some respects they are the most generalized of known arthropods, but if the arthropods have been developed from a segmented, wormlike progenitor provided with jointed legs, there is a vast gap between the trilobites and their vermiform ancestors. A trilobite is in no sense a primitive arthropod, and, notwithstanding all the claims that have been made in favor of a trilobite ancestry for the other arthropods, it is not probable that any other group of arthropods was derived directly from the trilobites. Any specialized form of animal produces only more specialized and diversified forms of its own kind.
Very soon after the beginning of the Cambrian the trilobites are accompanied in the geologic record by representatives of the Crustacea, the Eurypterida, and the Xiphosurida, but also there are various fossil arthropods found in the Cambrian rocks that cannot accurately be classified in these groups, but which appear to be related to them. Hence, it is to be supposed that the arthropods present in Cambrian and Ordovician periods represent lines of descent from common progenitors that lived far back in the immeasurable period before the Cambrian, and which, therefore, have little chance of ever being known. It is curious, however, that in the later Paleozoic rocks arachnids, myriapods, and insects appear as fully formed animals of their kinds, while there is nothing to show from what they came during the earlier Devonian, Ordovician, and Cambrian times, when trilobites, eurypterids, xiphosurids, and crustaceans were well preserved. Fossilization, however, depends largely on the presence of a hard skeleton, so that it is likely that animals first appear as fossils only when they have acquired a sufficiently resistant integument, and even then they must meet with favorable external conditions for fossilization, while, finally, fossils themselves may be utterly destroyed by subsequent metamorphosis of the containing rocks.
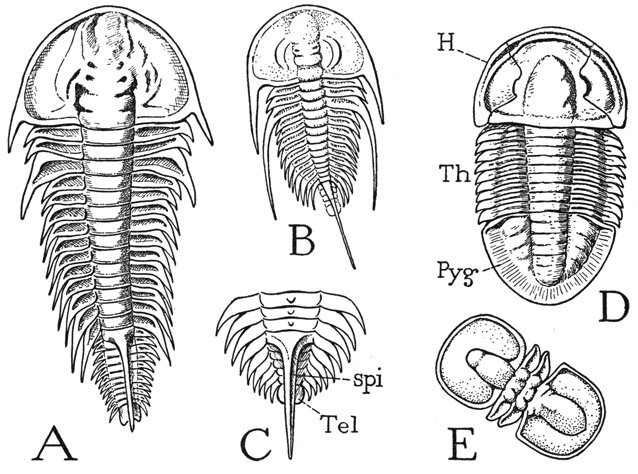
Fig. 1. Trilobita. Examples of trilobite forms.
A, Olenellus vermontanus Hall (from Walcott, 1910). B, Olenellus gilberti Meek (from Walcott, 1910). C, Schmidtiellus mickwitzi Schmidt (from Walcott, 1910). D, Asaphiscus wheeleri Meek (from Walcott, 1916). E, Peronopsis montis (Matthew) (from Walcott, 1908).
H, head, or prosoma; Pyg, pygidium; spi, spine; Tel, telson; Th, thorax.
The form and structure of the trilobites (fig. 2) show clearly that in these ancient animals the fundamental arthropod organization was already fully developed and had attained a specific type of specialization. Only in the lack of differentiation in the postoral appendages are the trilobites generalized, but their specialization is of a relatively simple kind that fitted them for life on the ocean bottom in shallow water along the shore, where most of them lived probably in the manner of the modern Limulus, though some species are thought to have been pelagic, or even deepwater inhabitants. Lacking jaws or grasping organs of any kind other than the legs, the trilobites could not have obtained active prey by raptatory methods, and it has been thought that probably they were mud feeders, but animals with long filamentous antennae can hardly be supposed to have made a practice of burrowing in mud or sand. In some species the projecting mesal ends of the coxae (fig. 2 B) are armed with spines, which fact suggests that the coxal lobes had some use in the obtaining of food; Raymond (1920) says, “The primary function of these spiny lobes of the coxa was doubtless the gathering and preparation of food, and carrying it to the mouth by passing it forward from one to the next.” On the other hand, since the coxal lobes do not meet in the middle line, and the spines are not well developed in all species, Störmer (1944) thinks it unlikely that they functioned as jaws. In any case, whatever may have been the food of the trilobites, and however they obtained it, the great numbers of the animals would indicate that they had an ample food supply within their reach. Certainly worms of various kinds as well as other soft-bodied creatures living in the mud or sand along the ocean shores were abundant in both Cambrian and Pre-Cambrian times. It would seem, in fact, that a trilobite should be quite fit to live under modern conditions, and paleontologists have no positive evidence to account for their early extinction.
General Structure of a Trilobite
Since few perfectly preserved specimens of trilobites are known, it is not possible to give a full description of the trilobite structure in any one species, but inasmuch as the details of structure have been carefully studied in different species, we can reconstruct diagrammatically, as given in figure 2, the general form and make-up of a typical representative of the group.
The body of a simple trilobite of usual form is oval and dorsoventrally flattened (figs. 1 D, 2 A); it is divided transversely into three parts known as the head (H), the thorax (Th), and the pygidium (Pyg). The thorax and pygidium together, however, may be said to constitute the body as distinguished from the head, but, since the so-called head bears the first four pairs of legs, the terms prosoma and opisthosoma are preferable names for the two principal parts of the animal. The prosomatic head is unsegmented in the adult, though it may show evidence of coalesced segments; the thorax is completely segmented; and the pygidium is clearly composed of united segments. In the earlier trilobites the pygidial region is fully segmented (fig. 1 A, B, C). Extending lengthwise usually through the three parts is a rounded, median elevation, which is flanked by wide, flattened or decurved lateral areas. On the head the median elevation is known as the glabella (fig. 2 A, Glb), the lateral parts as the cheeks, or genae; on the body the median elevation is termed the axis, or rachis (Rac), the lateral parts the pleurae (Plr). These terms and others to be introduced later are those current in trilobite taxonomy, and the parts they connote have no necessary relation to those similarly named in other arthropod groups. The name trilobite is derived from the apparent lengthwise division of the animal into three parts.
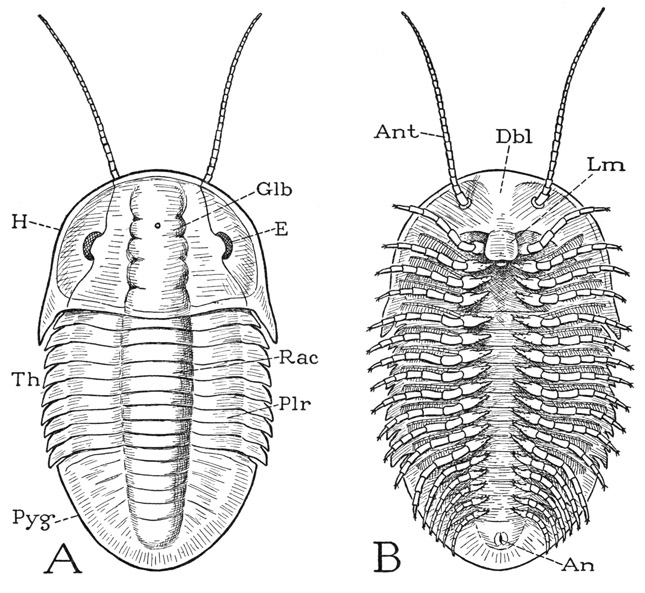
Fig. 2. Trilobita. Diagrams showing the structure of a generalized trilobite.
A, dorsal. B, ventral.
An, anus; Ant, antenna; Dbl, doublure; E, compound eye; Glb, glabella; H, head or prosoma; Lm, labrum or hypostome; Plr, pleura (not the pleuron of other arthropods); Pyg, pygidium; Rac, rachis or axis; Th, thorax.
Though the trilobite as seen from above (fig. 2 A) looks like no other animal with which we are familiar, the undersurface (B) at once shows that the trilobite is an arthropod; on each side is a long row of jointed legs, and from the anterior part of the head there projects forward a pair of slender, filamentous antennae (Ant). Behind the bases of the antennae and extended over the mouth is a median lobe (Lm), such as all arthropods have and which is commonly known as the labrum, though students of trilobites have generally called it the hypostome. The mouth presumably is covered by the labrum, and behind it is a small metastomal lobe. Of the legs, the first four pairs pertain to the head, the others to the thorax and pygidium, there being a pair for each body segment except the last. It will be noted that the legs are attached to the body at each side of a narrow median space, and that the pleural areas of the dorsum (fig. 4 A, D) are inflected on the undersurface to the bases of the legs. The inflected ventral surfaces laterad of the leg bases constitute the doublure (Dbl), which includes also the inflected undersurface of the head (figs. 2 B, 3 K).
A cross section through the thorax of a trilobite (fig. 4 A) clearly shows that the principal body cavity of the animal is in the median part, or rachis (Rac), which is strongly convex on the back, and that the so-called pleurae are merely flat hollow extensions of the body segments over the appendages. Since various other flattened arthropods have a similar structure, the “three-lobed” character is not distinctive of the trilobites. The special feature of the trilobites is the uniform, leglike structure of all the appendages except the antennae, in which respect the trilobites are more generalized than any modern arthropods, since in all of the latter at least some of the appendages are modified and specialized for purposes other than that of locomotion; the trilobites apparently did not even have appendages that served specifically as jaws. Details of the leg structure will be described in a later section.
The Head, or Prosoma
The head section, or prosoma, of a typical adult trilobite (fig. 2 A, H) is usually somewhat semicircular in outline, with its rounded anterior and lateral margins produced posteriorly in a pair of large genal spines. Between the bases of the spines the head is directly attached to the body by its transverse posterior margin, without the intervention of a neck. The dorsal surface of the head, as already noted, generally presents a median, elevated glabellar area (Glb) and broad, lateral genal areas, but in many species the head is covered by a perfectly smooth, rounded, shieldlike plate. The features of the head given in the following description, and illustrated diagrammatically in figure 3 L, have been made out from a study of many specimens of more generalized trilobites; the beginning student, however, is likely to see little trace of them in ordinary museum examples, owing partly to the imperfection or corrosion of the specimens, but also to the fact that the characters themselves were suppressed in the later evolution of the trilobites.
The grooves that separate the glabella from the genae are known as the dorsal furrows (fig. 3 L, df). A subdivision of the glabella into five successive parts, of which the first is the frontal lobe (frl), may be indicated by lateral notches in the dorsal furrows, or by imperfect transverse grooves in the glabellar surface. Each genal area is divided lengthwise by a facial sulcus, or “suture,” before and behind the eye (afs, pfs), which separates it into a median part called the fixed cheek, or fixigene (Fg), and a lateral part distinguished as the free cheek, or libragene (Lg). The free cheeks are produced posteriorly into the genal spines (gspi). An ocular ridge (er) goes from each side of the frontal lobe to the eye. The entire median part of the head shield between the facial sulci, including the glabella and the fixed cheeks, is termed the cranidium (Crn). A marginal furrow (mf) surrounds the head inside a narrow border area (b). Most trilobites have a pair of large, lateral compound eyes (E) on the mesal parts of the free cheeks, and above each eye a protective palpebral lobe (pbl) on the edge of the fixed cheek. On the anterior part of the glabella in some species there is present a small tubercle, which has been regarded as a median eye, but its ocular nature is questionable. A complete terminology for all parts of the trilobite is given by Howell, Frederickson, Lochman, Raasch, and Rasetti (1947).
Inasmuch as the trilobite head carries the first four pairs of legs, it must include at least four primitive postoral somites, and the glabellar grooves of the adult evidently represent the primary intersegmental lines of the head segments. The best understanding of the adult head has been derived from studies, such as those of Raw (1925), Warburg (1925), Lalicker (1935), Störmer (1942, 1944), and others, on specimens of very young trilobites in successive stages of development. All writers are in close agreement as to the visible facts, though they differ somewhat in their interpretations.
The youngest-known developmental stage of a trilobite is a minute oval thing, from half a millimeter to a millimeter and a half in length, and is termed the protaspis (fig. 3 A, E). The major part of the protaspis represents only the head of the adult trilobite. On the back of the youngest specimens a median glabellar elevation is already differentiated from wide lateral genal areas and very soon becomes divided transversely into five primary subdivisions, of which the last four (I–IV) represent the somites of the four pairs of legs carried by the adult head; faint intersegmental lines may be seen extending laterally in the genal areas (E). The first glabellar subdivision of the protaspis includes the area of the frontal lobe of the adult (J, frl) and its lateral extensions curving posteriorly around the anterior margin, which eventually become the free cheeks bearing the compound eyes. This anterior, or acronal, section of the larva may be termed the acron (A, Acr), though the term has been used with various other applications. Its underfolded anterior part forms the doublure of the head, on which the antennae and labrum of the adult are situated (fig. 2 B).
The preoral antennae of the trilobites are clearly the first antennae, or antennules, of other arthropods. The four postoral pairs of legs on the head, then, should reasonably be supposed to correspond with the first four postantennular appendages of other arthropods, which primarily arise behind the mouth, these appendages being the chelicerae, the pedipalps, and the first two pairs of legs in the Chelicerata, or the second antennae, the mandibles, and the two pairs of maxillae in the Mandibulata. If the acron of the trilobite larva is a single segment, the trilobite head is, therefore, composed of five primary somites, but the frontal lobe is sometimes divided into two parts. According to Störmer (1942), in the fourth protaspis stage of Olenus gibbosus (fig. 3 H) there appears in the sides of the frontal lobe a pair of pits, which eventually run together in a transverse groove, so that the frontal lobe becomes divided secondarily in development into two segments, which Störmer designates an antennal (i.e., antennular) segment and a preantennal segment. Henriksen (1926) and other writers, however, have contended that the posterior division of the frontal lobe is the segment of a suppressed pair of appendages representing the second antennae of Crustacea, and that the four persistent appendages are to be identified with the mandibles, the two pairs of maxillae, and the first maxillipeds. This contention that the trilobites have lost a pair of appendages, of which there is no concrete evidence, seems to be purely presumptive. “Such conclusions,” Störmer asserts, “have apparently been too much influenced by the current opinion of a crustacean nature of the trilobites.” In other words, a lost “second antenna” has been arbitrarily injected in order to make the trilobites conform with their supposed crustacean descendants. However, the trilobite, as it is, conforms with either the Chelicerata or the Crustacea on the assumption that its first legs represent the chelicerae of the former and the second antennae of the latter.
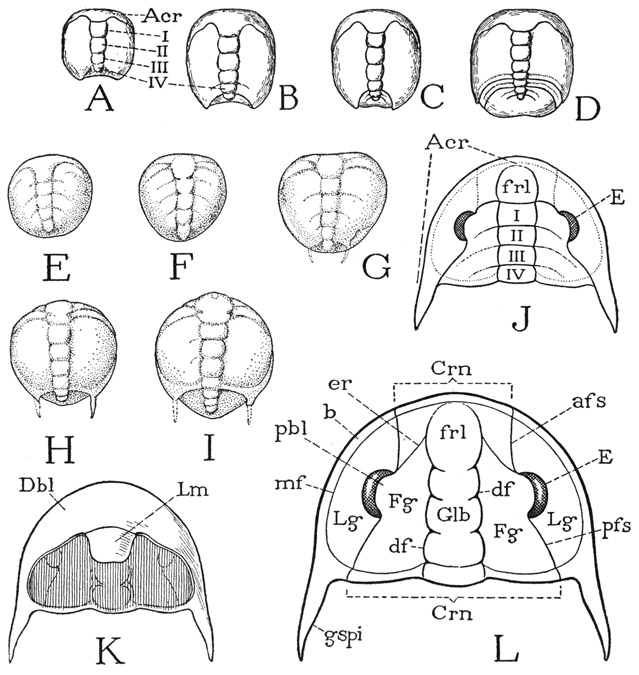
Fig. 3. Trilobita. Postembryonic development and the structure of the adult head, or prosoma.
A–D, developmental stages of Liostracus linnarssoni Brogger (from Warburg, 1925). E–I, developmental stages of Olenus gibbosus (Wahlenberg) (from Störmer, 1942). J, diagram of structure of mature trilobite head, dorsal. K, diagram of undersurface of head. L, diagram of dorsal surface of head, with parts named according to Howell, Frederickson, Lochman, Raasch, and Rasetti (1947).
Acr, acron, anterior unsegmented part of larva; afs, anterior facial sulcus; b, border area; Crn, cranidium; df, dorsal furrow; E, compound eye; er, ocular ridge; Fg, fixed cheek or fixigene; frl, frontal lobe; Glb, glabella; gspi, genal spine; Lg, free cheek or libragene; mf, marginal furrow; pbl, palpebral lobe; pfs, posterior facial sulcus; I–IV, primary postacronal segments of larva.
From the development of the protaspis it may be deduced that the pattern of the segmental composition of the adult trilobite head must be approximately that shown diagrammatically at J of figure 3. The oculoantennal part of the head, bearing the eyes dorsally and the antennae ventrally, which for convenience is here called the acron, has extended posteriorly on the sides from the frontal lobe, forming the regions of the free cheeks ending in the genal spines. The median part of the head, including the postfrontal part of the glabella and the fixed cheeks, is the region of the four postoral legbearing somites, but the intersegmental lines become obliterated in the adult except for remnants of them on the glabella.
The Body, or Opisthosoma
Though the five primary segments of the young protaspis all go into the formation of the adult head, there is a very small region behind the last head segment that is destined to generate the body. In the developing protaspis a stage is soon reached when a new segment appears behind the last head segment (fig. 3 B, G), and this segment is followed by others successively formed between it and the extending end of the body (C, D, H, I). These segments are the beginning of the series of body segments. The pygidial and thoracic segments, therefore, are secondary somites generated in the usual teloblastic manner from a subterminal zone of growth, just as are the secondary somites of the polychaetes and of those modern arthropods that have an anamorphic postembryonic development.
The Thorax— The thoracic segments of the adult trilobite are usually all of similar size and shape, but as between different species they are highly variable in number, since there may be from two to forty or more, though the usual number is perhaps between seven or eight and fifteen. The segments undoubtedly were movable on each other and connected by infolded membranous conjunctivae; probably they were hinged by articulations between the tergal plates, as are the abdominal segments of the crayfish, so as to allow up and down movements, though evidently there could have been little movement in a transverse direction. Many trilobites are found rolled ventrally upon themselves in the manner of certain modern isopods. The elevated median parts of the thoracic terga are distinctly demarked from the long, flattened or decurved pleural lobes on the sides, but, as already noted in cross section (fig. 4 A), the pleural lobes are merely lateral expansions of the body extending out beyond the leg bases. A cross section of the trilobite thorax resembles a cross section of Limulus (fig. 7 A), and in each animal the tergal plate, or carapace, must be interpreted as including not only the dorsal integument, but also the doublure (Dbl) on the undersurface to the bases of the legs. The projecting ends of the pleural lobes of the trilobites generally form a row of pleural spines, which may be uniform in size and shape or varied; in some forms particular spines are greatly elongate (fig. 1 B), in others all may be long and slender. The parts called “pleura” by students of trilobites are not to be identified strictly with the parts so termed in other arthropods. The true sternal area of the trilobite is the narrow ventral space between the leg bases and appears to have been a relatively soft integument.
The Pygidium— The pygidium is composed of a number of segments that are not fully differentiated as they are generated in development, but since in some of the earlier trilobites the body is completely segmented (fig. 1 A), it is probable that the pygidium has been formed in the course of evolution by a secondary union of the posterior body segments. The typical pygidium (figs. 1 D, 2 A, Pyg) is a large, smoothly rounded plate equal to the head in size, or sometimes larger than the head. It contains an extension of the rachis, and the pleural areas are marked by indistinct intersegmental lines. The pygidial appendages become successively smaller than those of the thorax (fig. 2 B). The anus (An) is situated ventrally in the apical segment of the pygidium, which, therefore, is to be regarded as the telson. In forms in which the pygidial region is segmented, one or more long median spines may project from the dorsum of the anterior pygidial segments (fig. 1 A, B, C). In one genus at least, the terminal segment bears a pair of long, multiarticulate tail filaments. It has been suggested that the pygidium was used as a swimming organ in the manner of the abdomen and “tail fan” of a crayfish.
The Appendages
The appendages of the trilobites are now fairly well known in a number of species; they include the antennae and the legs. In all the arthropods the antennae, or antennules, appear to be organs of a nature different from that of the other appendages, since in their normal development they never have the form or structure of ambulatory limbs, while, as the trilobites themselves attest, the other appendages, regardless of their form in modern arthropods, undoubtedly were all primarily walking legs. If the antennae were ever evolved from leglike appendages, they must have completed their transformation long before the time of the oldest-known trilobites.
The Antennae— The long, slender, tapering antenna of the trilobite is divided into a large number of short rings, and in appearance resembles the antenna of an orthopteroid insect, or the flagellum of a crustacean antenna, in which the rings are mere annulations without muscles, and therefore are not true segments, as are the musculated subdivisions in the antennae of the myriapods and the entognathous apterygote insects. The antennae of the adult trilobite arise from the doublure of the head anterior to the mouth (fig. 2 B, Ant), and are evidently the homologues of the antennules of Crustacea, though they are not branched, and of the antennae of the myriapods and insects.
The Legs— The legs of the trilobites are typical, uniramous, segmented arthropod limbs, with long, pinnate epipodites arising from their bases (fig. 4 A). Since each leg has seven clearly marked segments, the segmentation was formerly thought to correspond with that of a crustacean leg, and the epipodite was regarded as an exopodite, three small claws on the end of the segment being treated as mere apical spines. More recently, Störmer (1944) has shown that there is at the base of the leg a small ring by which the appendage is attached to the body, and this proximal ring Störmer regards as the true basal segment of the limb, which he calls a “precoxa,” or “subcoxa” (B, Prcx), the large segment following being the coxa. A petrified specimen, however, can hardly be supposed to give conclusive evidence that a basal ring of the appendage is a true segment in the sense of having been an independently movable section of the limb when the animal was alive, and it seems very improbable that the body muscles that moved the leg as a whole should have been attached on such a relatively small piece. In modern arthropods the basal muscles of an appendage are always attached on the coxa, unless the coxa itself has become immovable, but the base of the coxa is often set off as a proximal ring, or basicoxite, by an external groove that forms internally a strengthening ridge, or shelf, for the muscle attachments, and the basal ring of the coxa might superficially resemble a small segment. That the “precoxa” of the trilobite leg is a real limb segment, therefore, is open to question.
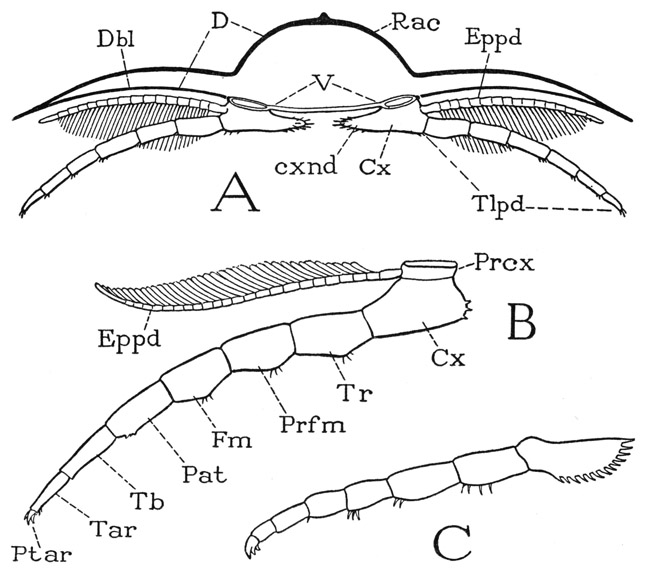
Fig. 4. Trilobita. Sectional structure of the body and segmentation of the legs.
A, diagrammatic cross section of a trilobite, showing attachment of the legs. B, a trilobite leg, with interpretation of segments according to Störmer (1944). C, leg of Neolenus serratus (Rominger) (from Raymond, 1920).
Cx, coxa; cxnd, coxal endite; D, dorsum; Dbl, doublure of dorsum; Eppd, epipodite; Fm, femur; Pat, patella; Prcx, precoxa; Prfm, prefemur; Ptar, pretarsus; Rac, rachis; Tb, tibia; Tar, tarsus; Tlpd, telopodite; Tr, trochanter; V, venter.
The rest of the leg beyond the subcoxal ring is divided into seven cylindrical sections (fig. 4 B), and there is no reason for supposing that these parts were not movable on each other, though, lacking any knowledge of the leg musculature, we cannot be so sure they were all true segments. The seventh apparent segment bears the three apical spines or claws, and Störmer, with good reason, has identified the claws as constituting the true end segment of the limb, since a similar three-clawed endpiece of the legs in other arthropods is always found to be a reduced, clawlike, apical segment provided with its own muscles and armed with a pair of lateral claws. A figure of an appendage of Neolenus serratus (fig. 4 C) given by Raymond (1920) leaves little doubt of the segmental nature of the apical part of the limb, though Raymond himself describes the claws as “little spines” on the end of the “dactylopodite.” The median claw in modern arthropods, however, is the true dactylopodite, of which the lateral claws are secondary outgrowths.
Discounting the segmental nature of the precoxa, then, the trilobite leg appears to be an eight-segmented limb. If so, the segments are to be identified as Störmer has named them (fig. 4 B): coxa (coxopodite), trochanter (basipodite), prefemur (ischiopodite), femur (meropodite), patella, tibia (carpopodite), tarsus (propodite), and pretarsus (dactylopodite), the names in parentheses being those used in carcinology. An eight-segmented limb depends on the coincidental occurrence of a patella and of two segments in the trochanteral region; it is not common in modern arthropods, but is present in the Pycnogonida and in some of the legs of the Solpugida among the arachnids. The patella is a segment characteristic of the legs of Arachnida and Pycnogonida and is not found in those of mandibulate arthropods. A tibia and tarsus normally follow the patella, or the femur if there is no patella; but it must be noted that the tarsus is prone to subdivision, particularly into two parts, which simulate segments, but are not interconnected by muscles. With regard to the trilobite leg, therefore, it might be questioned if the apparent tibia and tarsus are not two tarsal subsegments and the supposed patella the true tibia. There must, in other words, be some uncertainty as to the identity of limb segments where the musculature cannot be known. Yet the trilobite leg appears to be an eight-segmented appendage, not counting the subcoxal ring, and, if so, it is a truly generalized arthropod limb in that it contains all the segments present in the legs of any modern arthropod, but it particularly resembles the arachnid leg in the possession of a patella.
The long pinnate or plumose lateral branches arising from the bases of the trilobite legs (fig. 4 A, B, Eppd) are of particular interest. The earlier students of trilobite limbs regarded these branches as “exopodites,” and from this interpretation they drew the conclusion that the trilobite leg is a biramous appendage, and therefore closely relates the trilobites to the Crustacea. Störmer (1933), however, showed that the supposed exopodite arises, as he then thought, on the coxa and not on the basipodite (first trochanter) as does the crustacean exopodite, from which fact it obviously is an epipodite and not an exopodite; later (1939) he assigned it to the precoxal ring (fig. 4 B). Furthermore, Störmer gives good reasons for regarding the organs in question as gills. The epipodites are covered by the pleural lobes of the body segments, and, if they are not branchiae, the trilobites have no evident organs of respiration. In any case, the idea that the crustacean biramous type of limb represents the primitive and fundamental structure of arthropod appendages is not supported by the trilobites. The crustacean exopodite is always borne by the basipodite, and is a special feature of the Crustacea, though not present on all their appendages.