THE EURYPTERIDA
THE eurypterids were aquatic animals that lived during the Paleozoic era from the Cambrian into the Permian, but they attained their greatest development in the Silurian and Devonian periods. They are known also as the Gigantostraca because some of them attained the great length of six or eight feet, though most species are from six to 15 inches in length. The remains of the earlier forms are found in marine deposits along ancient shores or in lagoons, but later in the Devonian and Carboniferous periods the animals became fresh-water inhabitants.
The body of a typical eurypterid (fig. 13 A) is elongate, broad in front, tapering posteriorly, and usually ends in a large tail spine. As in the trilobites and the xiphosurids, the body is divided into an unsegmented prosoma and an opisthosoma, but the opisthosoma is always fully segmented. The prosoma carries six pairs of appendages (B) corresponding with the chelicerae and legs of Limulus, and therefore contains at least six segments. The opisthosoma in dorsal view (A) presents 12 tergal plates, showing that the entire body of the animal is composed of 18 postoral segments, not counting the tail spine. On the undersurface (B), however, there is evidence of only 11 segments in the opisthosoma (VIII–XVIII), suggesting that the sternal part of the seventh body segment is either obliterated or united with the prosoma. The eurypterid differs from the xiphosurid in that the tergum of the seventh segment is free from the prosoma (A, VII).
The prosoma is covered dorsally by a carapace of variable shape, either quadrate (fig. 13 D), rounded (figs. 13 A, J; 14 B, C), or triangular (fig. 14 A), but always of simpler structure than that of the xiphosurids. It bears a pair of lateral compound eyes (fig. 13 A, E) apparently similar in structure to those of Limulus, though in some species having an anterior position (fig. 14 A, C). Medially on the carapace is a pair of simple dorsal ocelli (fig. 13 A, dO). On the ventral surface of the prosoma (fig. 13 B) the limb bases occupy a greater area than in Limulus, because the prosoma is much less expanded and the doublure is therefore relatively narrow. The first of the appendages are a pair of three-segmented chelicerae, which are usually very small (B, G, Chl), but in one genus they are long arms (fig. 14 C) with large, strongly toothed chelae (E). The following appendages, or legs, are generally of simple form (fig. 13 G), increasing in size from before backward, and also in the number of segments, but in some species they are of diversified form and size (D, E). The usually very short first legs (G) have seven segments, counting the terminal spine as the apical segment, or pretarsus. The next two legs are successively a little longer and have eight segments each. The still longer fourth legs have nine apparent segments, as do also the much longer and broader last legs, which in most species have a paddlelike shape suggestive of their having been used as swimming organs.
It is difficult to account for the presence of nine segments in the last two pairs of eurypterid legs. The basal segment of these legs (fig. 13 H), as of the other legs, is undoubtedly the coxa (Cx), and the apical segment the pretarsus (Ptar). As in the case of the trilobite leg, however, it is impossible to be sure of the identity of the intervening segments without knowing the musculature. The penultimate and antepenultimate “segments” might be subdivisions of the tarsus, as they are in many other arthropods, including the scorpions, but in the eurypterid they appear to be true, independently movable segments. Suspicion, therefore, falls on the two small rings at the base of the trochanteral region of the leg, which may be supposed to be secondary subdivisions of the first trochanter (1Tr), since they are not present in the first three legs. In the lack of positive evidence, then, the other segments in the last two pairs of legs may be interpreted as the second trochanter, or prefemur (2Tr), the femur (Fm), the patella (Pat), the tibia (Tb), the tarsus (Tar), and the pretarsus (Ptar). In species having slender hind legs (fig. 14 B, D) the segmentation is the same as in the paddle-shaped legs. Between the coxae of the last legs is a large oval or elongate plate called the metastome (fig. 13 B, G, mpl). It has the appearance of being a sternal plate, perhaps the sternum of the seventh body segment pushed forward between the bases of the legs of the sixth segment, but some writers regard it as representing the united appendages of the seventh segment corresponding with the similarly placed chilaria of Limulus. Whatever the plate may be, it is not to be confused with the so-called “metastome” of Crustacea, which is an entirely different structure.
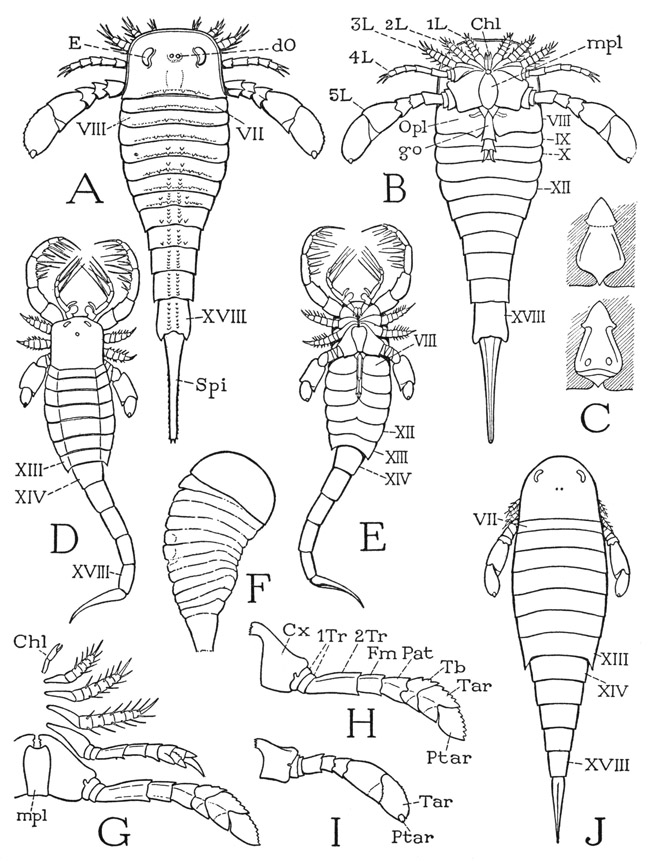
Fig. 13. Eurypterida. (A, B, F–I, outlines from Clarke and Ruedemann, 1912; C, from Störmer, 1936; D, E, J, from Störmer, 1934.)
A, Eurypterus remipes Dekay, dorsal. B, diagram of ventral surface of a eurypterid. C, Pterygotus rhenaniae Jaekel, opercular appendage of the broad type (female), ventral and dorsal. D, Mixopterus kiaeri Störmer, dorsal. E, same, ventral. F, Strabops thatcheri Beecher, upper Carboniferous merostome. G, Dolichopterus macrochirus Hall, left appendages and “metastome,” ventral. H, same, hind leg. I, Eurypterus remipes Dekay, hind leg. J, Hughmilleria norvegica (Kiaer), dorsal.
Roman numerals indicate body segments beginning with cheliceral segment. Chi, chelicera; Cx, coxa; dO, dorsal ocellus; E, compound eye; Fm, femur; go, genital organ; L, leg; mpl, metastomal plate; Opl, operculum; Pat, patella; Ptar, pretarsus; Spi, tail spine; Tar, tarsus; Tb, tibia; 1Tr, first trochanter; 2Tr, second trochanter.
The mouth of the eurypterid lies between the radiating bases of the prosomatic appendages and thus has a central position on the underside of the prosoma, as in the Xiphosurida. In a depression between the mouth and the metastomal plate is a lobe termed the endostoma. Some investigators claim that there is to be seen on the doublure in front of the chelicerae a small median organ, which is thought to represent the “ventral eye” of Limulus.
Of the 11 segmental divisions of the undersurface of the opisthosoma (fig. 13 B), the first corresponds with the eighth tergum of the dorsum (A). The venter of segment VIII is covered by a pair of plates (B, Opl) separated medially by a deep cleft from which projects a median appendage (go) of different form in the two sexes. By comparison with Limulus the two lateral plates would appear to represent the two halves of the operculum of the eighth segment, and the median organ the united telopodites of the opercular appendages. The two types of structure presented by the median organ of the eurypterids are designated A and B. Though both are highly variable, type A is generally a long slender structure (B, go), segmented in its distal part, while type B is short and broad, and not so distinctly segmented (C). It has usually been assumed that type A pertains to the female, and type B to the male, but Störmer (1936), from studies on Pterygotus rhenaniae von Overath, concludes that the reverse is more probably the truth. On the dorsal side of the base of the more slender organ he finds evidence of a median aperture, which he takes to be the opening of the male genital outlet duct; on the corresponding part of the broad organ (C), however, are two oval apertures that Störmer assumes to be the outlets of paired oviducts of the female.
The ventral surfaces of the next four segments of the opisthosoma are covered by broad sternal plates (fig. 13 B, IX–XII), which are said to be separate from the corresponding tergal plates. The part of the opisthosoma composed of the first six segments, segments VII to XII inclusive, therefore, is termed the mesosoma, and the following part, composed of segments XIII to XVIII, in which the segments are continuously sclerotized rings, is called the metasoma. In many species, however, the opisthosoma is abruptly narrowed between segments XIII and XIV (figs. 13 D, E, J; 14 A), so that the last five segments form a taillike part of the body, giving such eurypterids their characteristic scorpionlike appearance. The apical spine of the opisthosoma is usually straight and tapering in the eurypterids (figs. 13 A, B, J; 14 B) as in Limulus, but in the tailed species it may be curved and sharp-pointed (figs. 13 D, E; 14 A), suggesting the sting of a scorpion. In some forms, however, the apical structure is broad and fan-shaped (fig. 14 C).
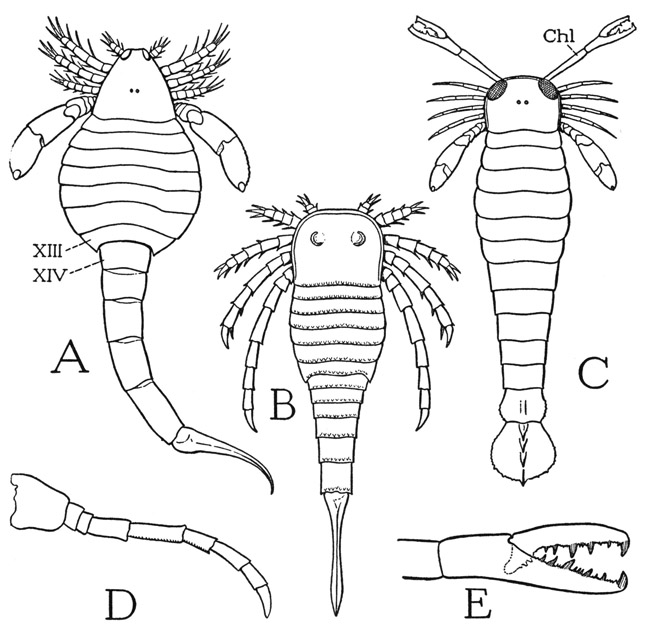
Fig. 14. Eurypterida. (A–D, outlines from Clarke and Ruedemann, 1912; E, from Störmer, 1936.)
A, Eusarcus scorpionis Grote and Pitt. B, Stylonurus longicaudata Clk. & Rued. C, Pterygotus buffaloensis Pohlman. D, Stylonurus longicaudatus Clk. & Rued., hind leg. E, Pterygotus rhenaniae Jaekel, reconstructed chelicera.
Lettering explained under figure 13.
In one respect the eurypterids differ conspicuously from the xiphosurids, which is the absence of exposed branchial organs on the opisthosoma. On the concealed dorsal surfaces of the opercular plates of the eighth segment, and on the inflected upper surfaces of the next four underlapping sternal plates, however, are paired structures that are interpreted as gills. In the Eurypterida, therefore, the five pairs of supposed branchiae occur on segments VIII to XII; in modern Xiphosurida the five pairs of gills are on segments IX to XIII.
The eurypterids undoubtedly must be in some way related to the xiphosurids. In the less elaborate development of the prosoma and in the segmentation of the opisthosoma they appear to be more generalized than the modern Limulus, but, as we have seen (fig. 8 E, F), the ancient fossil xiphosurids had a segmented opisthosoma. On the other hand, the scorpionlike form of some of the eurypterids has given rise to theories of relationship between the eurypterids and the scorpions. Probably the majority of zoologists with any opinion on the subject would regard the eurypterids as the ancestors of the scorpions, and therefore of the Arachnida in general, of which the scorpion is a member. It is certain, however, as Versluys and Demoll (1920) have emphatically asserted, that the scorpion is not a primitive arachnid; so these authors contend that the relationship is the other way around, namely, that the eurypterids and through them the xiphosurids have been evolved from scorpions, fossil scorpions being known from Silurian deposits. According to this theory the arachnids came from some independent source. Quite inconsistent with the scorpion ancestor theory, however, is the evident fact that the scorpionlike eurypterids are specialized forms, and that the earliest-known merostomes, from which the later eurypterids and modern xiphosurids must have descended, have no resemblance to scorpions, either ancient or modern. There is the Cambrian fossil Strabops (fig. 13 F), for example, which looks like a much-simplified eurypterid. Though paleontologists do not agree as to whether Strabops is a eurypterid or a xiphosurid, or whether it has 11 or 12 opisthosomatic segments, it is undoubtedly a primitive merostome, and has little to suggest that its ancestor was a scorpion. Moreover, historically the earliest eurypterids long antedate the first known scorpion. Neither theory of merostome-scorpion relationships, therefore, seems to be tenable. In the fourth chapter it will be shown that the scorpions differ in some important respects from the merostomes, and cannot be regarded even as primitive arachnids.