THE PAUROPODA
THE members of this group are minute, elongate, soft-bodied arthropods of the myriapod type of structure (fig. 70 A, B), but because of their relatively few legs, usually nine pairs in the adult stage, they have been named pauropods (Lubbock, 1868). A pauropod of average size is about a millimeter in length, but some species are only half as long, and others reach a length of nearly 2 mm. Probably owing to their small size, the pauropods have no circulatory system and no tracheae or other differentiated organs of respiration. They live in moist places under logs and stones, on the ground among decaying leaves, and in the soil to a depth of several inches. The feeding habits of the pauropods are not well known, but their food has been thought to be humus and decaying plant and animal tissue. Starling (1944) says that mold fungi were observed to be the usual food of Pauropus carolinensis and that a “correlation appears to exist between the optimum temperature for mold growth in general and high incidence of pauropod population.” He gives reasons for believing that pauropods, where abundant, regardless of their small size, play a significant part in soil formation.
A typical adult pauropod (fig. 70 B) has a relatively small, conical head and an elongate body of 12 segments, counting as segments the first and the last body divisions, which are known respectively as the collum (Col) and the pygidium (Pyg). Statements by other writers as to the number of segments may vary, because some do not include the pygidium as a segment and some exclude both the collum and the pygidium, but such differences are merely a matter of definition for a “segment.”
The number of legs in an adult pauropod, except in one known species, is invariably nine pairs, the first pair being on the second body segment, the last on the tenth (fig. 70 B). Preceding the pygidium is a legless segment, and the pygidium itself never bears appendages. A ten-legged pauropod, Decapauropus (F), said to have 13 body segments, has been described by Rémy (1931). In this form the usual eleventh segment appears to be partly subdivided ventrally into two parts, the anterior of which carries the extra pair of legs; but it might be suspected that the apparent division is superficial and that Decapauropus differs from other genera only in having legs on the eleventh segment.
The body segmentation of most pauropods appears to be different on the dorsal and ventral surfaces (fig. 70 B, F). In the specimen shown at A all the segments between the collum and the pygidium appear to be fully and equally developed, but generally, as seen in the genus Pauropus (B), there are only six tergal plates on the dorsum, though there are ten corresponding segmental divisions of the venter. This condition suggests the diplopod type of segmentation, but in the extended pauropod the narrowed dorsal arcs of segments 4, 6, 8, and 10 are exposed between the tergal plates of the other segments. Hansen (1902) has noted that live specimens of Pauropodidae “are able to elongate their body or to shorten it by contraction in a very high degree.” Each of the tergal plates except the first bears a pair of long, slender tactile setae.
The first segment, or collum, of the pauropod body (fig. 70 B) is quite different from the collum of a diplopod; instead of being a large tergal plate, it forms but an inconspicuous fold dorsally behind the head and is much expanded below. It bears ventrally a pair of small papillae (H, g), which have commonly been regarded as vestigial legs, but according to Tiegs (1947) they neither develop in the manner of limbs nor have the structure of legs, and appear rather to be comparable to the exsertile vesicles of Symphyla and apterygote insects.
The pauropod legs are all six-segmented (fig. 70 I), including the small pretarsus (Ptar). Except in the legs of the first and the last pairs, the tarsus of the adult (Tar) is divided into two subsegments. Writers who do not regard the pretarsus as a segment, state that the leg is either “six-segmented” or “five-segmented” according as the tarsus is divided or not. That the “segmentation” of the tarsus is not a division into true segments is shown by the fact that no muscles are attached on the distal part, and none arises in the proximal part. The entire tarsus is traversed in the usual manner by the tendon of the flexor muscle of the pretarsus (flptar), the branches of which arise in the tibia (Tb) and the femur (Fm). The musculature thus identifies the distal segments of the leg, and there is only one segment (Tr) in the trochanteral region between the coxa and the short femur; the leg, therefore, is six-segmented in all cases, regardless of the subsegmentation of the tarsus. The structure of the pretarsus is variable; there is a median claw and usually a ventral lobe, or empodium, but one or two accessory lateral claws may be present (fig. 70 J), and the claws themselves may be padlike on their undersurfaces.
The pauropod head has a unique feature in the presence of a pair of large, smooth, clear, oval areas on the sides (fig. 70 D, E, e) that have the appearance of eyes and were formerly supposed to be eyes. The sublying tissue, however, has nothing of the structure characteristic of light receptors, and the organs are now given the meaningless name of “pseudoculi,” their function being unknown. It is shown by Tiegs (1947) that the convex, cornealike cuticle of a pseudoculus is separated by an intervening space from a flat layer of large epidermal cells beneath it, which are innervated from the protocephalon. Tiegs suggests that the organs might be responsive to vibrations, but that, since the subcuticular space is apparently filled with liquid, the organs must be supposed to respond, by contact, to vibrations of solid objects rather than to air vibrations.
Characteristic of the pauropods are the relatively large, branched antennae (fig. 70 D). Each antenna consists of a basal stalk, which is four-segmented in the adult stage and of two apical branches, one dorsal, the other ventral. The dorsal branch ends in a single slender multiarticulate flagellum; the ventral branch bears two shorter flagella and, arising between their bases, a club-shaped or globular appendage termed the globulus (f), supposed to be a special sense organ of some kind. The segments of the antennal stalk and the bases of the branches are individually musculated, but the flagella have no muscles.
The pauropod mouth parts include only a pair of mandibles (fig. 70 H, Md) and an underlip structure (Mx, mxS), the latter, said to be composed of the appendages and sternum of the first maxillary segment, appears, therefore, to be comparable with the diplopod gnathochilarium. The pauropod mandibles, however, have no likeness to the jaws of a diplopod; the gnathal lobes are solid extensions from the elongate, tapering bases (G), which, instead of being fully exposed on the sides of the cranium, are deeply sunken into the head (E, Md), as are the mandibles of a chilopod. Insofar as the pauropod mandibles are solid, single-piece appendages they resemble in particular the mandibles of a geophilid chilopod. In Pauropus, according to Tiegs (1947), the tapering basal part of each mandible is connected by a ligament (E, d) to the head apodeme (hAp) of the same side and by a lateral ligament (not shown in the figure) to the cranial wall just behind the pseudoculus. This lateral ligament is suggestive of the slender rod connecting the end of the chilopod mandible with the cranial margin (fig. 59 B, mdr), which will be met with again in the Protura and Collembola. The ventrally concave distal ends of the pauropod mandibles are provided with combs of delicate curved blades (fig. 70 H), and together with a median elevation of the epipharyngeal surface, Tiegs says, they form the upper wall of a tubular food passage above the grooved floor of the preoral cavity. The mandibular musculature is described by Tiegs as consisting of protractor, retractor, depressor, and adductor muscles, the last attached on the head apodemes.
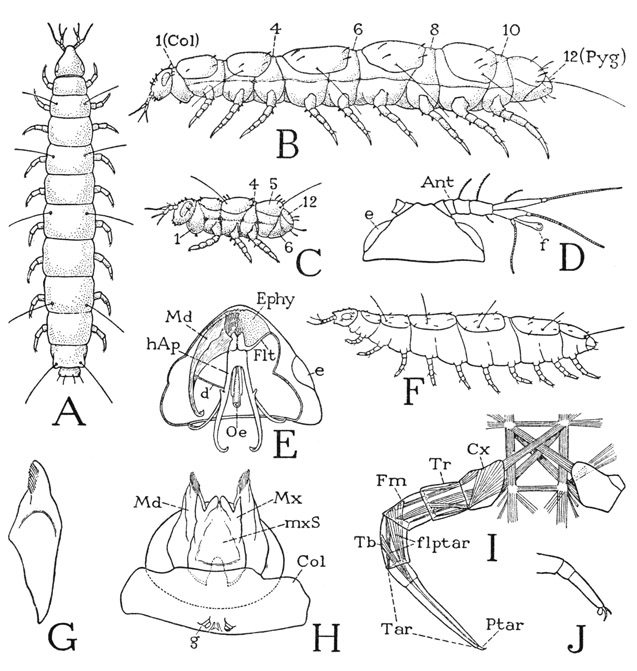
Fig. 70. Pauropoda. (B, C, E from Tiegs, 1947; D, G, H, I from Silvestri, 1902.)
A, a pauropod (unidentified) with regular segmentation on the dorsum. B, Pauropus silvaticus Tiegs, fully extended adult. C, same, first instar. D, Stylo-pauropus pubescens Hansen, head and right antenna, dorsal. E, Pauropus silvaticus Tiegs, underside of head, with right mandible in place and head apodemes. F, Decapauropus cuenoti Rémy (from Rémy, 1931). G, Allopauropus brevisetus Silv., mandible. H, same, head and collum segment, ventral. I, same, segmentation and musculature of a leg. J, a pauropod leg with accessory pretarsal claw and empodium.
Ant, antenna; Col, collum (first body segment); Cx, coxa; d, ligament from mandible to head apodeme; e, “pseudoculus”; Ephy, epipharyngeal surface; f, “globulus” of antenna; flptar, flexor muscle of pretarsus; Flt, fultural sclerite; Fm, femur; g, ventral papillae of collum segment; hAp, head apodeme; Md, mandible; Mx, maxilla; mxS, maxillary sternum; Oe, oesophagus; Ptar, pretarsus; Pyg pygidium; Tar, tarsus; Tb, tibia; Tr, trochanter.
Associated with the mandibles is a suspensory apparatus of the same nature as the hypopharyngeal fulturae of the chilopods. It consists of a pair of slender, transverse, premandibular sclerites in the ventral head wall attached laterally on the cranial margins (fig. 70 E, Flt). From the enlarged mesal ends of the sclerites, as described by Tiegs in Pauropus, arises a pair of large head apodemes (hAp). The bases of the apodemes unite in a sclerotic ring around the oesophagus (Oe); the distal parts are long, two-branched arms, of which the mesal pair extend back into the collum segment, while the lateral branches curve dorsally and are attached on the occipital margin of the head.
The postmandibular component of the pauropod mouth parts (fig. 70 H), representing the gnathochilarium of the diplopods, is said by Silvestri (1902), Tiegs (1947), and other investigators to include the first maxillary appendages (Mx) and a triangular median plate (mxS), which is the sternum of the first maxillary segment. The parts, however, are much less unified than in the diplopods. In Allopauropus (H), according to Silvestri, the maxillae are free from the median plate only at their distal ends. On the other hand, Hansen (1930) says that in Stylopauropus the maxillae are so sharply separated from the median plate that there can be no question of union between them. Tiegs, in his study of Pauropus silvaticus, makes no specific statement as to the degree of union of the maxillae with the sternal sclerite, but from his illustration (text fig. 2 B) it would appear that the three parts are quite separate. In any case, there is little doubt that the maxillae and the plate between them in the pauropods are the elements that are united in the diplopod gnathochilarium. The pair of median apical lobes, called “galeae” by Silvestri, are said by Tiegs to arise within the preoral cavity from the mandibular segment, “and seem therefore to be the equivalent of the superlinguae.”
The pauropods unquestionably are more primitive in their general organization than are the diplopods; the most important difference between the two groups is in the structure of the mandibles, but otherwise a close relationship is indicated by the segmental composition of the head, the primarily ventral position of the gonads, and the opening of the genital ducts on the third body segment. The development of the pauropod head is said by Tiegs (1947) to show that the adult head contains only one maxillary segment, the appendages and sternum of which form the gnathochilarium. The next segment, representing the second maxillary, or labial, segment of most other arthropods, becomes the first body segment, termed the collum segment. The ovaries of the pauropods remain throughout life between the alimentary canal and the nerve cord, and their ducts discharge through a single median pore into a depression of the venter on the third body segment between the bases of the second legs. A sperm receptacle opens from the dorsal wall of the genital depression. In the male pauropod the testes are formed ventrally during embryonic development, but, as shown by Tiegs (1947), they later acquire secondarily a dorsal position above the alimentary canal. The two ejaculatory ducts open separately through a pair of penes on the third segment mesad of the coxae of the legs. The exit genital ducts of the pauropods, Tiegs says, are not persisting coelomoducts, as in most other arthropods, but are epidermal ingrowths, which fact suggests that the functional ducts may not be the primary ducts, but secondary outlets connecting with the gonads.
The pauropods are anamorphic in their postembryonic development, and, as with the diplopods, the young pauropod when first exposed by rupture of the eggshell is still a quiescent embryo invested in a cuticular covering in which it completes its embryonic development. In this condition the embryo of Pauropus amicus is said by Harrison (1914) to remain for three days; then the investing cuticle splits lengthwise on the back, and the young, active, six-legged pauropod of the first instar becomes free. The young of Pauropus silvaticus, according to Tiegs (1947), has six body segments in front of the pygidium (fig. 70 C). Tergal plates are present on segments 2, 3, and 5, and the legs are carried by segments 2, 3, and 4. Starling (1944), however, gives only five body segments, exclusive of the pygidium, for the six-legged young instar of Pauropus carolinensis; the last segment he says is the definitive eleventh segment. There are four immature instars, during which the number of body segments increases and extra legs are added, but different observers are not entirely in accord as to the order of succession of the newly formed segments. According to Starling (1944), the prepygidial segment is a permanent segment from the beginning and becomes the definitive eleventh segment, while the new segments are generated in front of it. Silvestri (1902) says that the formation of new segments takes place between the last segment and the one preceding it, but since elsewhere he does not enumerate the pygidium as a body segment, his ultimo segmento may refer to the eleventh. The idea of a permanent preanal segment, however, is said by Tiegs (1947) to be an error, the new segments being generated between the eleventh segment and the anal segment, or pygidium.