THE DIPLOPODA
THE diplopods are distinguished from all other arthropods by the fact that most of their body segments are “double” and, as implied in the group name, carry each two pairs of legs. The double nature of the segments is evident not only from their two pairs of legs but also in the presence of two pairs of ganglionic centers in the ventral nerve cord, two pairs of lateral pouches of the male gonads, and two pairs of cardiac ostia and alary muscles. The double segments are termed diplosomites. Since it would seem most probable that the diplopods have been derived from ancestors having single segments, it may be supposed that the diplosomites represent two primitive segments united; in their ontogenetic development, as shown by Pflugfelder (1932), the double segments are formed from two consecutive sets of somite rudiments that are not differentiated into separate segments. It is not clear what advantage the diplopods have gained from having two pairs of legs on each functional segment, or, otherwise stated, in having the segments united in pairs. By comparison with the agile chilopods, they are sluggish creatures, their gait is a leisurely crawl, and they make no effort to escape capture. Their curious organization, however, gives the diplopods a distinctive place among the arthropods.
In certain features of the head and the mandibles the diplopods are evidently related to the chilopods, while, on the other hand, there is little doubt that they are allied to the pauropods and the symphylids. The four groups, therefore, have generally been classed together as the Myriapoda. The Diplopoda, Pauropoda, and Symphyla, however, differ from the Chilopoda in that the genital openings are near the anterior end of the body, instead of at the posterior end. They are hence distinguished as the Progoneata, or progoneate myriapods, in contrast to the opisthogoneate chilopods. It is possible, however, that the progoneate condition is not primary, since Tiegs (1940) has shown that there is evidence in the ontogenetic development of the symphylids of a primitive posterior connection of the genital ducts with the exterior. The definitive anterior exit ducts, he says (1947), in both Pauropoda and Symphyla are not persisting coelomoducts but ectodermal ingrowths.
The class Diplopoda includes a large number of species, which taxonomists divide into three principal groups: the Pselaphognatha, the Opisthandria, and the Proterandria. The Pselaphognatha are minute forms comprised in the family Polyxenidae, having not more than 13 body segments, and are recognizable by the presence of large tufts of hair on the back and sides of the body. The Opisthandria are of larger size, and the males are characterized by having the last pair or the last two pairs of legs modified as clasping organs by which the male in copulation holds the female, while he places the sperm in her genital openings by means of his mandibles. The Proterandria include most of the more common types of diplopods; their distinguishing feature is the modification in the male of one pair or both pairs of legs, usually on the seventh body segment, to serve as the functional intromittent organs, termed gonopods, by which the sperm is transferred from the male genital openings on the third body segment to the receptacles of the female on the corresponding segment. The Proterandria are divided into four orders: the Polydesmoidea, the Nematophora, the Juliformia, and the Colobognatha. Representatives of the Polydesmoidea and the Juliformia will be described here as examples of the more familiar types of diplopods. For a full account of the structure and classification of the diplopods the student is referred to the comprehensive works of Silvestri (1903), Verhoeff (1926–1931), and Attems (1926b).
The diplopods feed principally on decaying vegetable matter, particularly on the dead leaves of certain kinds of trees and shrubs; fresh leaves and fruit they seldom eat, and carnivorous habits have been observed only in members of one family, which are said to attack earthworms and phalangids. The alimentary canal is a wide, simple tube extending straight through the body, and has a single pair of excretory Malpighian vessels arising from the anterior end of the proctodaeum. Salivary glands of the anterior part of the head open into the preoral cavity through the epipharyngeal wall, and glands in the anterior region of the body discharge on the base of the inner wall of the gnathochilarium.
A distinctive feature of the diplopods is the nature of the respiratory system. The breathing organs are bundles of tracheae given off from hollow sternal apodemes that are open to the exterior by apertures called spiracles, closely associated with the bases of the legs. Descriptions of the apodemes and tracheae of various diplopod genera will be found in papers by Effenberger (1907), Ziegler (1907), Krug (1907), Wernitzsch (1910), Reinecke (1910), Voges (1916), Ripper (1931), and Seifert (1932). In general, the tracheae are fine, unbranched tubes, with spiral taenidia, and do not unite with each other, but in the Polyxenidae and Glomeridae they are branched. The apodemes, which are variously developed, serve also for the attachment of the leg muscles, and usually the apodemes of the first three body segments have no tracheae. It seems probable that the apodemes were developed originally for muscle attachments; the spiracles of the diplopods evidently can have no relation to the lateral spiracles of the chilopods and insects.
In their mode of development the diplopods are anamorphic, as are the Scutigeromorpha and Lithobiomorpha among the chilopods. The young of most species leave the egg with only three pairs of legs, which are the first three pairs of the adult, and have not more than seven body segments. The definitive segmentation is acquired gradually at the subsequent moults, accompanied by an increase in the number of legs. The new segments are formed in a zone of growth at the posterior end of the body, which remains active throughout life. Inasmuch as adult diplopods of different groups differ much in the number of body segments, the anamorphic progression of segment formation is not the same in all of them. A detailed account of the manner of segment generation in the zone of growth is given by Pflugfelder (1932). The postembryonic development of Julus is described by Krug (1907), that of several species of Polydesmoidea by Miley (1927), Pflugfelder (1932), and Seifert (1932), and a general account of diplopod anamorphosis is given by Attems (1926b). Each successive stage of growth is accompanied by a moult, preliminary to which the young diplopod encloses itself in an earthen nest. At each moult the head capsule of the exuviae is disarticulated from the body, and ecdysis is accomplished by the animal’s crawling out of the old cuticle of the trunk. Miley (1927) observes that in Euryurus the shed cuticle is “split down the midventral line and also somewhat laterally, just above the articulation of the legs on one side or on both sides.”
The Head and the Mouth Parts
The diplopod head (fig. 63) is strongly convex above (D) and flattened beneath. Eyes are present in some forms (C, D) and absent in others (A). The antennae arise well back from the anterior margin, so that before them is a wide epistomal area (A, D, Epst), but the latter is in no way specifically separated from the rest of the cranial wall. With the margin of the epistome is united the narrow, dentate labrum (A, C, Lm). On the sides of the head are the large, convex bases of the mandibles (A, B, C, D, mdB), and the ventral surface of the head is covered by a broad flat appendage known as the gnathochilarium (B, D, Gnch). The antennae are seven-segmented appendages (A, Ant), as are the legs, and each segment is individually musculated. The eyes, when present, have superficially the appearance of compound eyes (C, D), but they are merely groups of simple eyes like those of the chilopods, as seen at F, G, and H on figure 69.
Postantennal organs of Tömösvary are present in most of the diplopods, except Juliformia and Colobognatha. In some forms the sensory cells lie beneath open pits or grooves; in others they are entirely covered by the cuticle. In the polydesmoids each organ is marked externally by an oval groove of the cuticle with a thickened central disc (fig. 63 A, OT); the sense cells lie beneath the groove and have no communication with the exterior.
Beneath the epistome (fig. 63 F, Epst) is a long palatal, or epipharyngeal, surface (Ephy) which extends posteriorly from the labrum (Lm) to the mouth (Mth) and forms the roof of a large preoral food cavity (PrC) closed ventrally by the gnathochilarium. At the base of the inner surface of the gnathochilarium is a group of flattened suboral lobes that constitute the hypopharynx (Hphy), which in the diplopods does not have the form of a projecting tonguelike organ, such as that of the chilopods and insects. The preoral cavity is occupied mostly by the gnathal lobes of the mandibles, which close against the hypopharynx. The epipharyngeal surface usually presents a transverse ridge behind the labrum (E) and a low median elevation.
The segmental composition of the diplopod head is a subject on which there is difference of opinion. The principal question is whether or not it contains a second maxillary segment. The anatomy of the adult head throws little light on the matter. On the back of the head is a narrow postoccipital arch (fig. 63 G, Poc), set off by a postoccipital sulcus (pos), which forms a large internal postoccipital ridge (poR) for the attachment of muscles from the body. The pattern of the structure is thus almost identical with that of the back of an orthopteroid insect head (fig. 78 B), which presumably contains the second maxillary segment. In the diplopod, however, the base of the mandible (fig. 63 B, D, G, mdB) comes back to the postoccipital sulcus, apparently leaving no room for more than one maxillary segment. The subject will be further discussed in connection with the gnathochilarium.
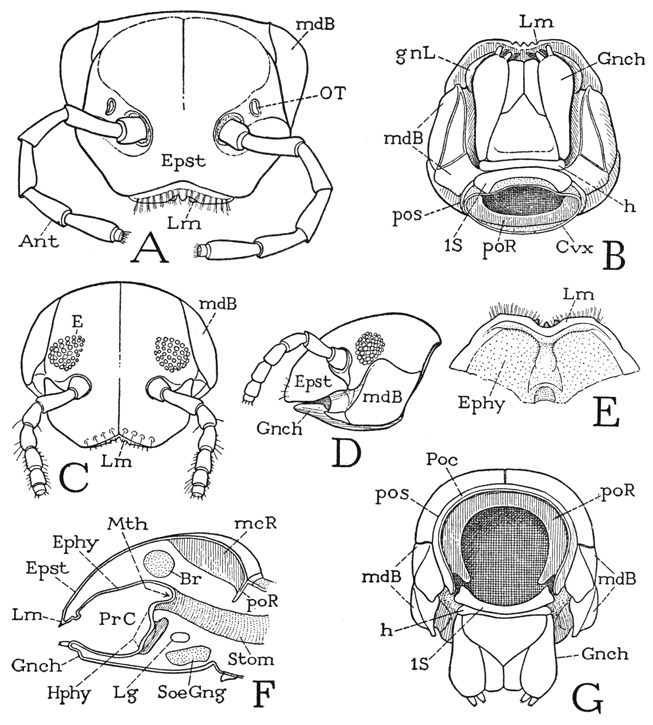
Fig. 63. Diplopoda—Polydesmoidea and Juliformia. The head.
A, Apheloria coriacea (Koch), head, anterior. B, Habrostrepus sp., head, ventral. C, Arctobolus marginatus (Say), head, anterior. D, same, head, lateral. E, Fontaria virginiensis (Drury), preoral epipharyngeal surface and labrum, ventral. F, same, diagrammatic longitudinal section of head. G, Habrostrepus sp., head, posterior.
For explanation of lettering see pages 248–249.
The mouth parts of the diplopod include the mandibles, the gnathochilarium, and the hypopharynx. The diplopod mandible (fig. 64 A, B) is the most remarkable type of jaw found among the arthropods. It consists of a large, convex basal part (mdB) and of a movable, independently musculated gnathal lobe (gnL) articulated on the base. As already noted, the basal part is set on the side of the head (fig. 63 D, mdB) between the margin of the cranium and the gnathochilarium. It consists usually of two distinct plates, one anterior, the other posterior, and the former may have secondary divisions. The mandibular base looks so much like a part of the head wall that it has been regarded as being the “pleuron,” or “ pleurites,” of the head; German writers commonly call it the Backen, or “cheek.” That the part in question, though little movable, is the true base of the mandible, however, is evident from the insertion, within its cavity (fig. 64 A), of huge muscles that clearly represent the ventral adductors of the mandibles of other arthropods. One or two groups of these muscles (D, 4b) go into each jaw from opposite ends of a thick, intergnathal ligament (Lg), and another posterior group (G, H, 4a) arises on the head apodeme (hAp) of the same side. A muscle inserted distally on the dorsal margin of the mandibular base arises posteriorly on the cranium.
The gnathal lobes of the mandibles (fig. 64 A, gnL) are articulated on the distal ends of the basal plates, from which they turn mesally within the preoral cavity, where they are mostly concealed from below (fig. 63 B, gnL) by the gnathochilarium. Each lobe is strongly but variously toothed at its apex (fig. 64 A, B, C) and has a rasping surface on the basal part of its mesal margin. The two opposed, independently movable lobes constitute the functional jaws of the diplopod. Each is provided with a flexor muscle of great size (B, 1) arising dorsally on the cranium and inserted by a thick, rigid tendon on its mesal basal angle. In some forms at least, a second, smaller muscle of the lobe arises in the distal part of the mandibular base. It is thus interesting to note that the musculature of the freely movable gnathal lobe of the diplopod mandible corresponds exactly with that of the flexible lobe of the chilopod mandible (fig. 59 E), suggesting that the diplopod jaw could have been developed from a mandible like that of the chilopods.
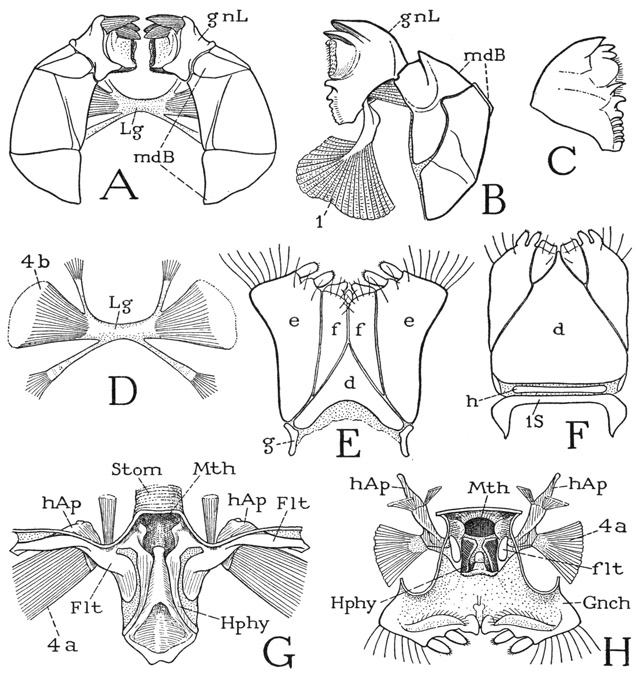
Fig. 64. Diplopoda—Polydesmoidea and Juliformia. The mouth parts.
A, Apheloria coriacea (Koch), mandibles, ventral. B, Arctobolus marginatus (Say), left mandible, with muscles of gnathal lobe, ventral. C, same, gnathal lobe of mandible, dorsal. D, Fontaria virginiensis (Drury), intermandibular ligament and muscles. E, Apheloria coriacea (Koch), gnathochilarium. F, Arctobolus marginatus (Say), gnathochilarium and first segmental sternum. G, same, hypopharynx and fultural sclerites (Flt) bearing head apodemes (hAp), dorsal. H, Apheloria coriacea (Koch), dorsal surface of gnathochilarium, with hypopharynx, showing reduced fulturae (flt) supporting long head apodemes.
For explanation of lettering see pages 248–249.
In describing the act of feeding by Strongylosoma pallipes Oliv., Seifert (1932) says the contraction of the muscles of the Backen (mandibular bases) pulls the latter mesally, and with them the gnathal lobes, which work independently by their own muscles. When this diplopod feeds on dry leaves, the food is first flooded with a secretion from the mouth that softens the surface between the leaf veins so that it is easily scraped or pulled off. The first three pairs of legs may serve as accessory feeding organs, small pieces of leaves being held by them and finally thrust between the mandibles and the gnathochilarium by the first pair.
The gnathochilarium is the functional underlip of the diplopod. It is attached by its base to the posterior part of the undersurface of the head (fig. 63 B, Gnch). The sclerotic ventral surface of the organ in most of the diplopods is divided by grooves into several characteristic parts (fig. 64 E, F), including a triangular median plate (d) known as the mentum, two lateral lobes (E, e, e) called the stipites, bearing each a pair of apical papillae, and a pair of median distal plates (f, f) termed the laminae linguales. In some species there is a narrow transverse sclerite, the prebasilare (F, h) intervening between the mentum and the first body sternum (1S), and at the sides, proximal to the stipites, are two small sclerites termed the cardines (E, g). The nomenclature of the parts of the gnathochilarium is based on the commonly held opinion that the organ is a composite of the usual first maxillae and the labium, or second maxillae. Students of the development of the gnathochilarium, however, are not in agreement as to its composition: some claim that it is formed by the union of both pairs of maxillae, others contend that it contains only the appendages and a sternal plate of the first maxillary segment, while still others assert that it is formed of the second maxillae alone, the first maxillae being suppressed during development. According to Silvestri (1903), the lines in the outer wall of the typical gnathochilarium are differentiated secondarily during embryonic development, and in the pselaphognathid Polyxenus the gnathochilarium is shown by Reinecke (1910) to be a simple structure without subdivisions.
The gnathochilarium is supported at its base on a ventral plate, commonly called the “hypostome,” which is the sternum of the first body segment (fig. 64 F, 1S). This segment, however, has been shown by Silvestri (1903, 1950) to be the second maxillary, or labial, segment of the chilopods and insects, and to have no appendages in the diplopods. The gnathochilarium, therefore, according to Silvestri, is a product of the union of the first maxillae, together with a sternal plate of their segment, and the diplopod head does not contain a second maxillary segment. We shall later see that, according to Tiegs (1947), the same is true of the pauropod head.
The hypopharynx of other arthropods is represented in the diplopods by the assemblage of flat lobes or folds of the integument situated in a depressed suboral area at the base of the inner wall of the gnathochilarium (fig. 64 G, H, Hphy). Typically there are a median lobe and two lateral lobes, which assume different shapes and proportions in different species. A supporting apparatus of the hypopharynx, consisting of a pair of transverse fultural sclerites attached laterally on the cranial margins, is fully developed in some of the diplopods, as in the spiroboloid shown at G (Flt). In the polydesmoid (H), however, the fulturae are reduced to small plates (flt) at the sides of the hypopharynx. In either case the sclerites support a pair of head apodemes (hAp) that give attachment to the second adductor muscles of the mandibles (4a). In the spiroboloid (G) the apodemes are short and thick; in the polydesmoid (H) they are elongate arms projecting posteriorly and dorsally in the head, with their apices attached to the central discs of the organs of Tömösvary (fig. 63 A, OT). Midway on each of these apodemes are attached two small muscles arising on the dorsal wall of the cranium, so that in this case the apodemes would appear to act as levers for lifting the hypopharynx. The presence of fultural sclerites of the hypopharynx bearing the head apodemes in Chilopoda, Diplopoda, and Pauropoda (see Tiegs, 1947) is a strong point in evidence of a relationship between these three myriapodous groups.
The Body Segments and Legs of a Polydesmoid Diplopod
The body of a polydesmoid diplopod (fig. 65 A) consists of 20 segments, as indicated by the number of back plates. The first segment behind the head and the last two segments are legless. The second, third, and fourth segments have each one pair of legs, the others bear each two pairs, including in the male the appendages of the first pair on the seventh segment, which are modified for reproductive purposes and become the gonopods. The consecutive segments are freely movable on each other, since they are united by wide, conjunctive membranes, but the conjunctives are ordinarily not visible, since normally the anterior part of each segment is deeply inserted into the posterior part of the segment preceding. Inasmuch as the double segment is the characteristic feature of the diplopod, it will be appropriate to understand first the structure of a diplosomite, preferably one from the middle part of the body.
Each diplosomite of Apheloria coriacea is a continuously sclerotized annulus with no sutural divisions between the tergal, pleural, and sternal areas (fig. 65 D, E). The segment, however, is demarked into an anterior ring and a posterior ring by an encircling groove a little before the middle. The anterior ring is termed the prozonite (Prz), the posterior ring the metazonite (Mtz). The dorsal surface of the metazonite is extended on each side in a broad lateral lobe (B, D, E); the long lateral surfaces of the segment between the tergal lobes and the bases of the legs (E, Pl) may be regarded as the pleura, or pleural areas, and the ventral wall carrying the legs the sternum (S). Since both pairs of legs are attached on the metazonite (D, C), it is evident that the two annular parts of the diplosomite do not represent the component segments. The prozonite has a smooth polished surface and fits into the metazonite of the preceding segment, thus permitting the characteristic ventral flexion or curvature of the body. Near the margin of each tergal lobe on most of the segments is a small pore (B, D, E, rp), which is the outlet of a repugnatorial gland; the pores are present on segments 5, 7, 9, 10, 12, 13, and 15 to 19 (figs. 65 A, 67 A). When the animal is irritated, the glands discharge a liquid with an acrid odor, said to contain free hydrocyanic acid, which is deadly to most other small arthropods in confined spaces.
On the undersurface of a diplosomite (fig. 65 C) it is seen that the two legs on each side arise close together and that those of the second pair are somewhat nearer the mid-line. In front of the outer angle of each leg base is a conspicuous oval pit (aSp, pSp). These pits lead into hollow cuticular ingrowths of the body wall (F, aAp, pAp) that serve both as apodemes for muscle attachments and as air passages for tracheae opening from them. The external openings of the apodemes, therefore, are the breathing apertures, or spiracles, of the diplopod; they are not simple holes, each orifice being guarded by a grating of branched processes that admits air but keeps out solid particles.
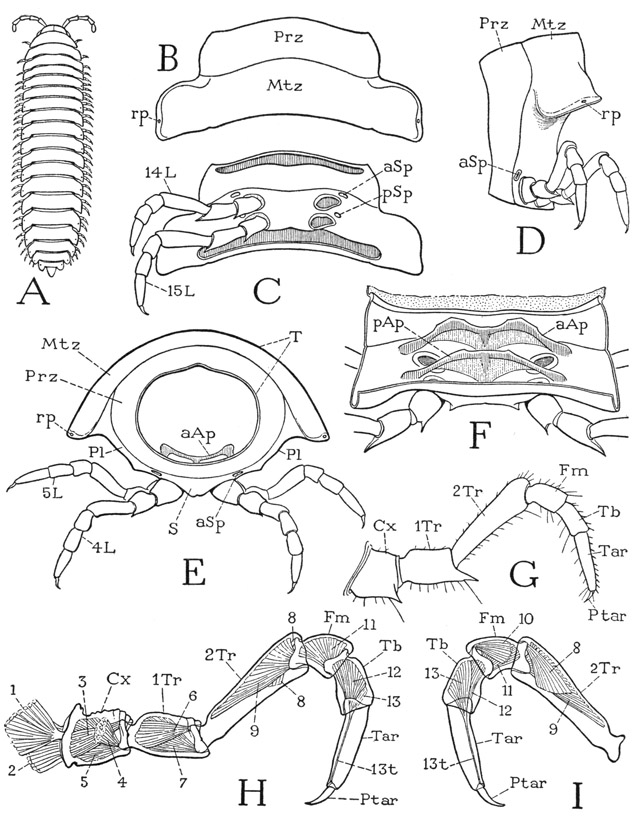
Fig. 65. Diplopoda—Polydesmoidea. Structure of body segments and the legs.
A, a typical polydesmoid diplopod. B, Apheloria coriacea (Koch), tergum of tenth segment. C, same, undersurface of tenth segment. D, same, lateral view of a diplosomite. E, same, fifth segment, anterior. F, same, inner surface of ventral wall of a diplosomite, showing ventral apodemes. G, same, a left leg, anterior. H, Euryurus sp., left leg and musculature, anterior. I, same, distal part of leg, posterior.
For explanation of lettering see pages 248–249.
An examination of the internal surface of the ventral wall of a diplosomite of Apheloria (fig. 65 F), when the muscles and other soft tissues have been removed by caustic, shows that it is crossed by two apodemal arches (aAp, pAp) between the opposite pairs of external pits. Each apodeme consists of a thick, arcuate anterior bar and a thin posterior lamella strengthened medially by a thickening from the marginal bar. The two apodemes are inclined forward, so that the posterior one partially overlaps the one in front. The lateral parts of the apodemal arms are hollow air chambers, from which numerous delicate tracheae arise in bundles (fig. 66 D, Tra). Since the apodemes give attachment to leg muscles and are present in the first three body segments, which have no tracheae, and inasmuch as many other arthropods have sternal apodemes for muscle attachment, it seems probable that the respiratory function of the diplopod apodemes is secondary.
The legs of Apheloria, as in most of the diplopods, are uniformly seven-segmented (fig. 65 G), and the tarsi are undivided, but the identity of the segments has been differently interpreted by different writers. Silvestri (1903) regarded the basal segment as a subcoxa, the second segment as the coxa, and the third as a trochanter. On the other hand, according to Attems (1926b), the first segment is the coxa, the second the prefemur, or trochanter, the third the femur, and the fourth a postfemur. Hansen (1930) calls the fourth segment the tibia and the fifth the cotibia. However, if we simply follow the usual nomenclature of the arthropod leg segments, beginning with the coxa, the segmentation of the diplopod leg will be as given at G and H of figure 65. The long third segment in the leg of Apheloria (G) or Euryurus (H) thus becomes the second trochanter, though it resembles the femur of an insect leg, but in other diplopods the leg segments may be subequal in length, and in the arthropods generally the relative length of a leg segment is no criterion of its identity. Between the coxa and the first trochanter there is sometimes present a small ring, but no muscles arise within it or are inserted on it, so that there is nothing to suggest that the ring represents a true segment.
The musculature of the diplopod leg is fairly simple. The limb as a whole turns forward and backward on the base of the coxa, and Silvestri (1903) has given a comparative account of the coxal muscles, all of which arise on the corresponding sternum and the sternal apodemes, but in some forms some of the muscles cross over from the apodeme of one side to the coxa of the opposite side. Each of the first four segments in Euryurus (fig. 65 H, I) following the coxa has levator and depressor muscles, but the long tarsus has only a depressor, or flexor (12). The simple, clawlike pretarsus (Ptar) has a single muscle, the usual flexor (13), arising in the tibia, with its fibers inserted on a long tendon (13t) traversing the tarsus from its attachment on the base of the pretarsus.
If we now turn to the anterior part of the body of Apheloria, it will be seen that several of the segments following the head differ from the diplosomites and, in various ways, from one another. The first back plate immediately behind the head (fig. 66 A) is known as the collar, or collum (Col). On the ventral surface below the collum is a small but well-developed sternal plate (F, 1S) bearing a pair of lateral apodemes (Ap); its extended anterior angles are connected with the posterior margin of the head by two crescent-shaped cervical sclerites (cvpl). The collum and this associated sternal plate will be here treated as representing a legless first segment of the body.
The first pair of legs (fig. 66 G, 1L) are suspended from the anterior margin of a second small sternal plate (2S) lying free in the ventral membranous integument behind the first sternum. This plate bears a pair of typical sternal apodemes (Ap) connected by a transverse bridge, but the apodemal pits lie posterior to the leg bases instead of anterior to them as in the diplosomites. Corresponding to this first leg-bearing sternum is a small tergal plate (B, 2T) behind the collum, which has large lateral lobes, from which descend normal pleura (C, Pl), but, instead of joining the sternum, the pleura are connected with each other ventrally by a narrow bridge behind the sternum.
The third body segment (fig. 66 H) carries the second pair of legs and is the segment of the genital openings in both sexes. The sternum is a distinct plate lying between the lower ends of the prozonite of the back plate, but the lower ends of the pleural areas of the metazonite are united in a narrow bridge behind the sternum. The sternum bears a pair of long tapering apodemal arms (Ap) connected by a transverse bar with a triangular median extension. There are no tracheae that have their origins in the first three segments, these segments and the head being tracheated from the fourth segment. The second pair of legs (H, 2L) are carried by the sternum of the third segment. In the male of Apheloria the coxa of each of these legs bears mesally a small genital papilla, or penis (Pen), through which a vas deferens (Vd) opens to the exterior after turning downward anterior to the apodemes.
In the female the venter of the third segment contains a pair of deep pockets lying behind and somewhat to the sides of the coxal bases. These pockets are closed posteriorly by a narrow poststernal bridge of the pleura; they contain the external female genitalia, which are known as the vulvae. When a vulva is protracted, or removed from its pouch (fig. 66 I), it is seen to consist of a short, thick, cylindrical basal stalk bearing a pair of soft, apical, valvelike lobes (vlv). Covering the lateral ends of these two transversely elongate vulval lobes is a smaller opercular lobe (opl) arising from the side of the stalk. Beneath the operculum is the large opening of the oviduct; it will be freely exposed when the operculum, which covers it like a lid, is lifted. A fourth, larger sclerotized lobe projects mesally from a fold of the pouch wall around the base of the vulva and forms a hoodlike covering over the vulval lobes when the latter are retracted. At the bottom of the depression between the vulval lobes (J) is a median groove that opens into a thick-walled, serpentine sperm receptacle.
The fourth body segment (fig. 66 D) carries the third legs and is the last segment with a single pair of appendages; otherwise it has the typical segmental structure. The tergum (4T) is produced into lateral lobes, below which the pleura converge ventrally and are continuous with the sternum; the coxal foramina are entirely enclosed in the ventral sclerotization (E). The apodemal pits of the fourth segment are the first pair of spiracles (Sp) and are particularly large; the tracheae (D, Tra) arising from the hollow lateral parts of the apodemes are distributed to the first four body segments and the head.
The next two segments, the fifth and the sixth, are typical diplosomites, each bearing two pairs of legs and containing two separate sternal apodemes.
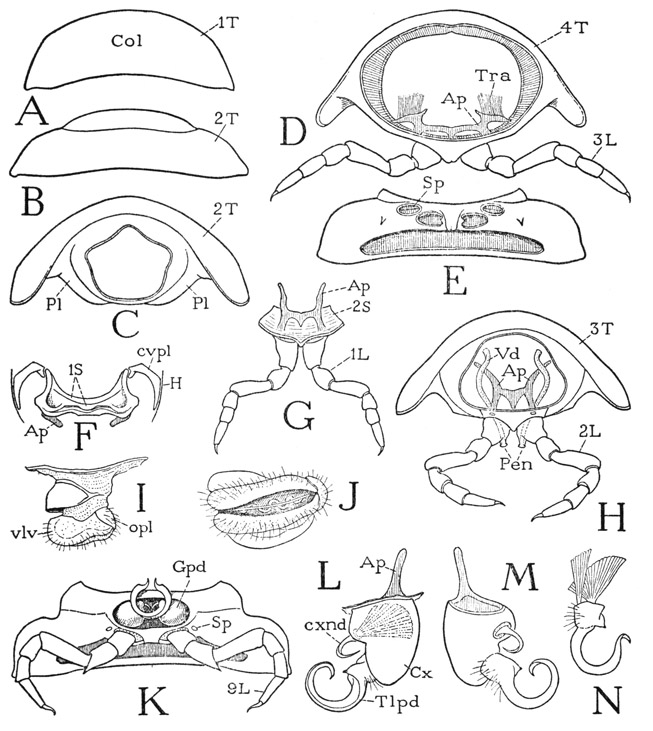
Fig. 66. Diplopoda—Polydesmoidea. Apheloria coriacea (Koch). Anterior body segments and gonopods.
A, first tergum, collum. B, second tergum. C, second segment, anterior, sternum (G, 2S) removed, pleura united ventrally behind sternum. D, fourth segment, posterior. E, same segment, ventral. F, first sternum, ventral. G, second sternum with first legs, anterior. H, third segment of male, with second legs, showing genital outlets on coxae, anterior. I, right vulva of third segment of female, posterior. J, same, ventral. K, seventh segment of male, ventral, with gonopods and legs. L, left gonopod, anterior. M, same gonopod, posterior. N, telopodite of gonopod, detached with muscles.
For explanation of lettering see pages 248–249.
The seventh body segment is of particular interest in the male because it is on this segment in most of the proterandrious diplopods that one pair or both pairs of legs are modified in structure to serve for transfer of the sperm from the genital openings on the third segment of the male to the sperm receptacles of the female. The modified appendages, as already noted, are known as gonopods. In Apheloria, as in other Polydesmoidea, only the first pair of appendages on the seventh segment are gonopods (fig. 66 K, Gpd), the second pair being ordinary seven-segmented legs (9L). In the female the same segment has the usual two pairs of legs. The gonopods of different species present a great variety of structure; their characters are much used by taxonomists for species identification.
The gonopods of Apheloria coriacea are sunken into a deep, transversely oval depression before the legs on the venter of the seventh segment (fig. 66 K, Gpd). Each gonopod (L) has a large, thick, conical basal segment (Cx), probably the coxa, from which an apodemal arm (Ap) projects into the body cavity. Distally, on its mesal surface, the coxa bears a long, sickle-shaped process, regarded as the telopodite (Tlpd), which tapers to a sharp, recurved point. Proximal to the base of the telopodite is a coxal endite in the form of a strongly bent hook (cxnd) set into an oval membranous area of the coxal wall (M). The telopodite is movable by antagonistic muscles (N) arising on the posterior wall of the coxa; the hook is provided with a single large fan-shaped muscle (L) arising on the anterior coxal wall. The mesal surface of the telopodite is almost flat, but it is traversed by a minute canal extending to the tip from a depression on its base. In the natural position (K) the telopodites of the opposite gonopods curve mesally and forward with their ends together or overlapping. Between them they enclose a cuplike space, the diameter of which is just sufficient to receive the two penes of the second legs (H, Pen), so that the apertures of the penes can be applied to the cavities at the inner ends of the telopodite canals, which are the sperm conduits of the intromittent apparatus. Copulation and the process of sperm transfer in two other polydesmoids, Strongylosoma pallipes and Polydesmus edentulus, are fully described by Seifert (1932). By bending the anterior part of the body downward, the male diplopod brings the penes on the second legs into contact with the gonopods on the seventh segment, and the sperm is discharged into the cavities on the bases of the telopodites, where it is mixed with a secretion from glands in the gonopods. During copulation, then, the sperm is discharged through the sperm canals of the gonopods into the sperm receptacles in the vulvae of the female, probably being activated by the copious secretion of the vulval lobes. That the telopodites and the coxal hooks also play an active role in copulation is indicated by their strong musculature.
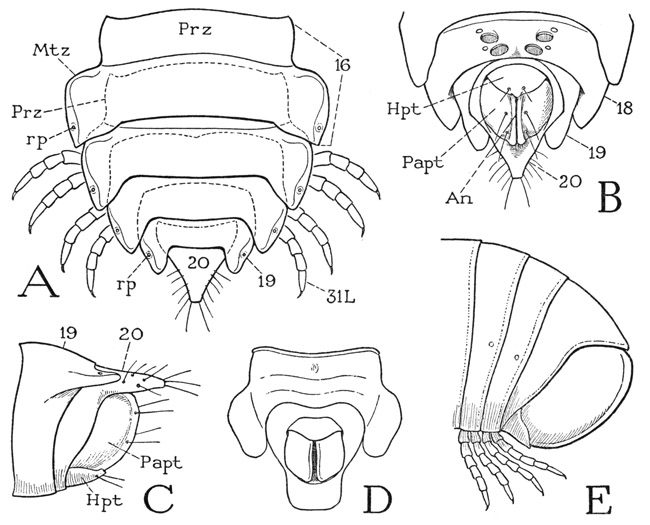
Fig. 67. Diplopoda—Polydesmoidea and Juliformia. Posterior body segments.
A, Apheloria coriacea (Koch), last five body segments, dorsal, with overlapped prozonites indicated by dotted lines. B, same, terminal segments, ventral. C, same, terminal segments, lateral. D, Euryurus sp., anal and preanal segments, ventral. E, Arctobolus marginatus (Say), terminal segments, lateral.
For explanation of lettering see pages 248–249.
According to Seifert (1932), the females of Strongylosoma pallipes lay their eggs in small round chambers constructed in the earth, the walls of which are glazed, apparently with a secretion from the anal glands. In each chamber 40 to 50 round, ivory-white eggs are deposited, after which the nest is carefully closed. Two or three nests may be made in succession by the same female to accommodate eggs ripening later in the ovaries. Other polydesmoids, however, may construct nests on objects above ground.
The next 11 segments of Apheloria are ordinary diplosomites and are essentially all alike in structure, but those beyond the sixteenth segment become successively narrower (fig. 67 A), and the body somewhat abruptly tapers posteriorly to a point. Segments 19 and 20 are legless (B, C). Segment 19 is a simple ring with relatively large tergal lobes, in which are the apertures (A, rp) of the last pair of repugnatorial glands. Segment 20 is produced dorsally into a triangular median lobe that forms the apical point of the body; its side margins bear a pair of valvelike lateral anal lobes, or paraprocts (B, C, Papt), and from its sternal arc projects a rounded subanal lobe, or hypoproct (Hpt). The anus lies in the vertical cleft between the lateral lobes. The twentieth segment, together with the anal lobes, probably represents the telson, since in the diplopods generally the zone of growth lies in the preceding segment.
The Body Segments of a Juliform Diplopod
The order Juliformia includes the cylindrical diplopods with long flexible bodies, numerous segments, and relatively small legs; when disturbed they coil themselves ventrally. Most of them have eyes (fig. 63 C, D), and there is a narrow sclerite, the “prebasilare,” between the gnathochilarium and the first body sternum (fig. 64 F, h). On the seventh segment of the male are usually two pairs of gonopods, but if only one pair is present there are no legs. The order includes three suborders: the Juloidea, the Spiroboloidea, and the Spirostreptomorpha. The following description is based mostly on the spiroboloid Arctobolus marginatus (Say), common in the eastern and southern parts of the United States.
The body of Arctobolus marginatus consists of about 50 segments. The segments beyond the second are short rings (fig. 68 A), somewhat wider above than below, and are but little inserted into each other; they are not differentiated into a prozonite and metazonite as in the polydesmoids, and the animal coils itself by a telescoping of the ventral parts of the segments rather than a separation of the dorsal arcs. The spiroboloids differ from other diplopods in that the fourth and fifth pairs of legs, as well as those preceding, are borne each on a separate segment (A, 4L, 5L). The first segment having two pairs of legs is the sixth, which carries the sixth and seventh legs. The next segment is that of the gonopods in the male and has two pairs of ordinary legs in the female.
The first body tergum is the collum (fig. 68 A, Col), a large, hoodlike plate extending over the back of the head as far as the eyes. The sternum of the collum segment (fig. 64 F, 1S) is a smaller and simpler plate than that of Apheloria and is more closely associated with the base of the gnathochilarium and the back of the head. The first legs hang from beneath the lower angles of the collum (fig. 68 A), but they are attached on a V-shaped second sternal plate (B, 2S) suspended in the membranous ventral integument below the collum, and for this ventral plate there is no corresponding tergal plate. The legs of the second pair (C, 2L) hang from a similar third sternum (3S), but this sternum is attached to the anterior margin of the second tergal plate (2T), which is expanded ventrally on the sides of the body (A), and the pleural areas of the segment are inflected mesally as a pair of flat triangular lobes (C, Pl), the points of which approach each other behind the coxae but do not unite. It is evident, therefore, that in this diplopod the second segment is incomplete in that it has no tergum, and that the actual second tergum (A, 2T) pertains to the third segment. The first two pairs of legs are closely associated with the head.
In the male of Arctobolus the vasa deferentia come together and open into a depression behind the coxal bases of the second pair of legs, but there are no penes. In some of the Juliformia, as in Habrostrepus (fig. 68 D), a pair of penes (Pen) arises posterior to the bases of the coxae, and in others, as in Parajulus (E), there may be only a single median penis. The first and second legs of Parajulus (1L, 2L) and related forms are modified in a very unusual manner.
The third, fourth, and fifth segments of Arctobolus are simple rings (fig. 68 A), each bearing a single pair of ordinary walking legs. The coxae of these legs (F) are produced into apical lobes. The sixth segment is the first diplosomite with two pairs of legs (A), and the seventh is the segment of the gonopods in the male. It will be noted that there are the same number of tergal plates (six) anterior to the gonopod segment in the spiroboloid as there are in the polydesmoid Apheloria, and also the same number of legs (seven), but that in the polydesmoid the fourth and fifth legs are carried by a single segment. On the other hand, the polydesmoid has a complete segment for the first legs, which segment in the spiroboloid is not represented by a tergal plate. While the number of apparent segments in the two forms is thus the same, the segments are not entirely identical.
The gonopods of Arctobolus marginatus are ordinarily concealed in a deep cavity on the venter of the seventh segment between a posterior sternal bridge and a transverse fold of the integument behind the preceding segment. When they are protracted, or removed (fig. 68 H), it is seen that an anterior V-shaped sternal plate (S) lies in front of the gonopods and is closely associated with them. The first gonopods (1Gpd) are attached to the ends of the sternal arms and clasp the bases of the second gonopods (2Gpd). Each first gonopod (I) is two-segmented; the basal segment, or coxa (Cx), is produced into a broad endite (cxnd); the distal segment, or telopodite (Tlpd), is an arm of irregular shape muscularly movable on the coxa. The second gonopod (J, K) is larger than the first, but its basal part is normally concealed in the sternal cavity. The coxa (Cx) runs out dorsally into an apodemal extension (Ap) and bears ventrally the telopodite (Tlpd), which is a large, thick arm with a bifid tip, turned mesally from its coxal articulation, strongly musculated, and deeply grooved on its dorsal surface for the reception of sperm from the third body segment. In the natural position (H) the telopodites of the second gonopods are clasped between the two branches of the first gonopods.
The gonopods are not in all proterandrious diplopods limited to the seventh segment. In the Nematophora the second legs of the sixth segment may be gonopods, or also the first or the second legs of the eighth segment. In the Colobognatha the gonopods are the second appendages of the seventh segment and the first appendages of the eighth segment, and in this group the gonopods resemble ordinary legs; in the genus Brachycyba, for example, each gonopod is merely a diminutive six-segmented leg.
The body segments following the segment of the gonopods in Arctobolus are all typical diplosomites and are almost circular in transverse outline (fig. 68 G). The segmental sterna are distinct plates (S) extending anterior to the bases of the first pair of legs; the coxae of the second pair are set into notches of the posterior sternal margin. Associated with the base of each leg is a spiracle. The spiracular apodemes in Arctobolus, however, though they give attachment to the leg muscles, are extremely small, thin, delicate, triangular, horizontally overlapping lamellae, similar to those of Julus as described by Krug (1907), Ziegler (1907), and Ripper (1931). The legs are more slender than those of Apheloria, but they have the same segmentation.
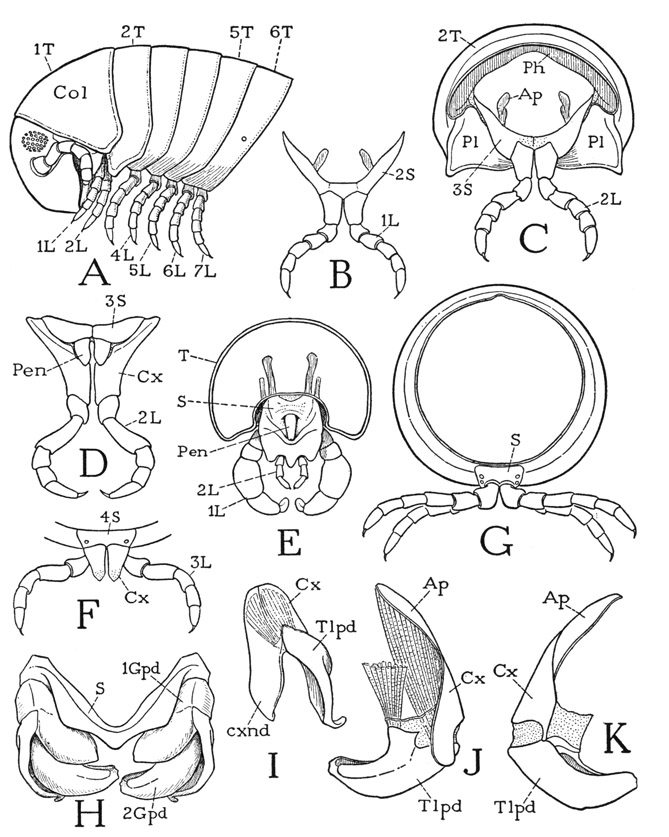
Fig. 68. Diplopoda—Juliformia. Body segments, legs, and gonopods.
A, Arctobolus marginatus (Say), head and anterior six body segments, lateral. B, same, first legs of male, anterior. C, same, second segment of male, anterior. D, Habrostrepus sp., male, second legs and supporting third sternum, paired penes arising at bases of coxae, posterior. E, Parajulus sp., male, first legs and segment of second legs with single median penis, posterior. F, Arctobolus marginatus (Say), sternum of fourth segment, with third legs, anterior. G, same, a diplosegment from middle part of body, anterior. H, same, gonopods and sternum of seventh segment, anterior. I, same, left first gonopod, anterolateral. J, same, left second gonopod, anterior. K, same, left second gonopod, posterior.
For explanation of lettering see pages 248–249.
The posterior end of the body of the spiroboloid is bluntly rounded (fig. 67 E). There is only one legless segment, which corresponds with the anal segment of Apheloria (C, 20) since it bears the lateral and ventral anal lobes. In the Nematophora the anal segment carries two or three pairs of slender, hollow spines that serve as outlets for silk-producing glands. The silk is discharged in the form of threads, and from this fact the Nematophora derive their name.
Eyes of Diplopoda and Chilopoda
The eyes of the diplopods and chilopods are in all cases lateral eyes; they are of particular interest in a comparative study of the optic organs of the arthropods because of their resemblance to the lateral eyes of certain apterygote insects and holometabolous insect larvae. Each eye is formed from a single layer of epidermal cells (fig. 69 A), which grows inward beneath the cuticular lens in the form of a cup (B, C). The outer cells of the cup (B, CgCls) retain the corneagenous function; the inner cells (SCls) become the sensory retinal elements. During the formative stage of the eye, and at each subsequent moult, the corneagenous cells extend in a continuous layer beneath the eye cuticle to form the lens, and after the completion of the lens they retract again to the periphery. The inner ends of the retinal cells directed toward the axis of the eyecup acquire light-sensitive striated borders (B, C) and may become elongated as terminal optic rods of the retinal cells (G, H). In some of the chilopods the retinal cells lose their uniformity of structure characteristic of the simpler types and become differentiated into a basal group of short central cells (D, cSCls) and an outer group of long peripheral cells (pSCl) converging beyond the basal cells. In this type of development the chilopod eye takes on the structure of the eyes of some holometabolous insect larvae.
The eyes of diplopods retain the simple cup-shaped type of retinal structure (fig. 69 F, G), in which the sensory cells are of approximately uniform size and have their receptive ends convergent along the axis of the eye. In some diplopods, as in Glomeris (F), the eyes are few in number and well separated, and each eye has a thick, strongly convex lens. In the Julidae, on the other hand, there are numerous closely contiguous eyes in each lateral group (G), having a common corneal covering (Cor) with a smooth outer surface. The inner surface of the cornea, however, is produced into thick lens processes (Ln) projecting individually into the retinal cups.
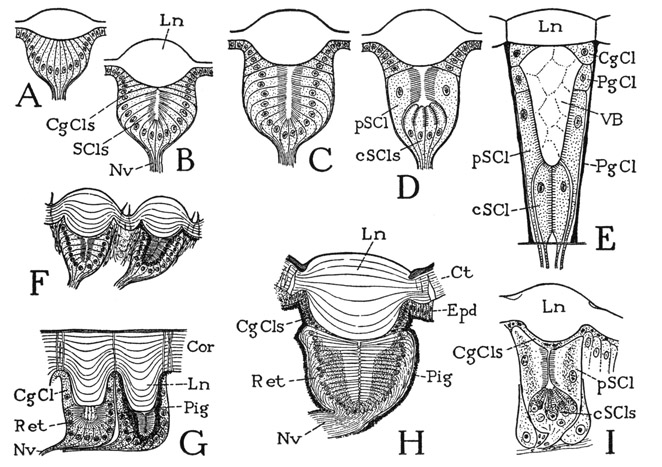
Fig. 69. Eyes of Chilopoda and Diplopoda.
A–D, diagrams of development of the eyecup. E, diagram of eye of Scutigera (based on Grenacher, 1880, Adensamer, 1894, and Hemenway, 1900). F, section of two adjacent eyes of Glomeris (from Grenacher, 1880). G, section of two eyecups of Julus (from Grenacher, 1880). H, section of eye of a scolopendrid, Heterostoma australicum (from Grenacher, 1880). I, section of eye of Lithobius (from Willem, 1892).
CgCls, cornagenous cells; Cor, cornea; cScls, central sense cells; Ct, cuticle; Epd, epidermis; Ln, lens; Nv, nerve; PgCl, pigment cell; Pig, pigment; pScl, peripheral sense cell; Ret, retina; SCls, sense cells, retina cells; VB, vitreous body.
Among the chilopods the Geophilomorpha are mostly without eyes; in the Scolopendromorpha the Scolopendridae have typically four small eyes on each side of the head, but the Cryptopidae are eyeless; the eyes of Lithobiomorpha have from 1 to 25 or 30 ocular units, those of Scutigeromorpha from 1 to 200. In the eye of a scolopendrid (fig. 69 H) the retinal cells (Ret) are of uniform elongate shape, and their distal receptive parts form cylindrical or polygonal optic rods directed horizontally toward the median axis of the eye-cup. In Lithobius (I) the retinal cells are differentiated into an inner globular group of central cells (cSCls), with optic rods convergent distally, and a single layer of large, distal peripheral cells (pSCl) disposed radially around the axis of the eye, with their receptive surfaces contingent beyond the central cells. The numerous eyes in a compact group on each side of the head of Scutigera (fig. 54 A, B) give the appearance of a pair of large compound eyes. Each individual eye, moreover, has the form of a tubular ommatidium (fig. 69 E) tapering toward the inner end and contains a large, conical vitreous body (VB) beneath the lens. The retinal part of the eye resembles that of Lithobius in that it consists of a distal set of long peripheral cells (pSCl), 10 to 12 in number according to Hemenway (1900), that embrace the sides of the vitreous body and of a proximal central group (cSCl) of 3 or 4 cells. The entire eye is surrounded by a sheath of pigment cells (PgCl). The nature of the vitreous body is not known, but the body is composed of a number of polygonal sections, in which, according to Adensamer (1894), nuclei may be seen in immature specimens. There can be little doubt that the compact group of simple eyes of Scutigera must function in the manner of an appositional compound eye, but the structural differences between the chilopod eyes and the compound eyes of crustaceans and insects are sufficient to show that there can be no relation between them. On the other hand, the structure of the eye of Scutigera is almost a replica of that of the eyes of dytiscid, trichopterous, and lepidopterous larvae among the insects (see Snodgrass, 1935, fig. 281).
Explanation of Lettering on Figures 63–68
aAp, anterior apodeme of a body segment.
An, anus.
Ant, antenna.
Ap, apodeme.
aSp, anterior spiracle of a segment.
Br, brain.
Col, collum.
cvpl, cervical plate.
Cvx, cervix, neck.
Cx, coxa.
cxnd, coxal endite.
d, “mentum” of gnathochilarium.
e, “stipes” of gnathochilarium.
E, eye.
Ephy, epipharynx.
Epst, epistome.
f, “lamina lingualis” of gnathochilarium.
flt, a much reduced fultural sclerite.
Flt, fultura, supporting sclerite of hypopharynx.
Fm, femur.
g, “cardo” of gnathochilarium.
Gnch, gnathochilarium.
gnL, gnathal lobe of mandible.
Gpd, gonopod; 1Gpd, 2Gpd, first and second gonopods.
h, “prebasilare” of gnathochilarium.
H, head.
hAp, head apodeme.
Hphy, hypopharynx.
Hpt, hypoproct.
L, leg.
Lg, intergnathal ligament.
Lm, labrum.
mcR, midcranial ridge.
mdB, base of mandible.
Mth, mouth.
Mtz, metazonite.
opl, opercular lobe of vulva.
OT, organ of Tömösvary.
pAp, posterior apodeme of a body segment.
Papt, paraproct.
Pen, penis.
Ph, phragma.
Pl, pleuron.
Poc, postocciput.
poR, postoccipital ridge.
pos, postoccipital sulcus.
PrC, preoral food cavity.
Prz, prozonite.
pSp, posterior spiracle of a segment.
Ptar, pretarsus.
rp, pore of repugnatorial gland.
S, sternum.
SoeGng, suboesophageal ganglion.
Sp, spiracle.
Stom, stomodaeum.
t, muscle tendon.
T, tergum.
Tar, tarsus.
Tb, tibia.
Tlpd, telopodite.
1Tr, first trochanter.
2Tr, second trochanter, prefemur.
Tra, tracheae.
Vd, vas deferens.
vlv, valvelike lobe of vulva.