THE SYMPHYLA
THE symphylans are small, slender, soft-bodied, progoneate, centipedelike arthropods with long antennae, 12 pairs of legs in the adult stage, and two tapering, unjointed appendages projecting from the rear end of the body (fig. 71 A). They feed mostly on vegetable matter and, in general, are found in moist places under stones and in rotting logs, but they live also in the ground, and one species at least, known as the “garden centipede,” becomes in some localities a serious pest of outdoor garden crops and of greenhouse plants.
In a zoological way the Symphyla have become of particular interest to entomologists because they have been much investigated as possible ancestors of the insects. Though in important features the symphylans more closely resemble the diplopods, they do have other characters that are found elsewhere only in the lower insects. In the structure of the mandibles, in the ventral position of the gonads, and particularly in the anterior position of the genital openings on the third body segment, the Symphyla are diplopodan; in the presence of two distinct pairs of maxillary appendages on the head and of eversible vesicles and styli on the venter of the body segments they seem to show some relation to the apterygote insects. Finally, however, as will be shown in the next chapter, the insects themselves have important features that could not have been derived from symphylan ancestors. In short, theoretical arthropod phylogeny becomes so complicated that we had best leave it to the phylogenists. The best presentation of the evidence adduced to support the symphylan claim to insect ancestry has been given by Imms (1936).
There are between 50 and 60 described species of Symphyla distributed throughout the world, classified by Attems (1926a) in three families, Scolopendrellidae, Scutigerellidae, and Geophilellidae. The most important recent papers on their structure, development, and life history are one by Michelbacher (1938) on the structure, life history, and economic aspect of the “garden centipede,” Scutigerella immaculata (Newport) and another by Tiegs (1940) on the structure and embryonic development of Hanseniella agilis Tiegs. Papers of lesser scope will be mentioned in the text. The following account is based mostly on Scutigerella immaculata, with comparative notes from Tiegs on Hanseniella agilis. Both species belong to the Scutigerellidae.
General Structure of the Body
Scutigerella immaculata, in common with other Symphyla, has 12 pairs of legs when it becomes an adult (fig. 71 A), but the dorsum of the body presents a series of 15 tergal plates. Three of the back plates, however, are duplicate terga of segments 3, 5, and 7, there being no corresponding ventral segmentation of the body. The nerve ganglia and the embryonic coelomic sacs are said to correspond in number with the legs. In the genus Geophilella there are as many as 22 tergal plates, but only the usual 12 pairs of legs. The segment numbered 13 in the figure of Scutigerella (fig. 71 A), known as the preanal segment, bears a pair of tapering, unjointed appendages (G, Spn), which are functionally spinnerets, since silk-forming glands open through them, but developmentally they arise in line with the leg rudiments and are therefore regarded as true segmental appendages. The preanal segment in most species carries also the last pair of legs, and thus might seem to be a double segment. In Scutigerella immaculata (G) there is a narrow but distinct circular fold (12) of the integument between the preanal segment (13) and the preceding segment (11), and the legs of the last pair (12L) appear to be attached to this fold rather than to the segment of the spinnerets. It is possible, therefore, that the fold represents the true twelfth segment, and the possibility is much increased by the fact that in a species of Symphylella (J), described by Tiegs (1940), there is in front of the pregenital segment (13) a well-developed extra segment (12) carrying the twelfth legs. In species with only 13 body segments, therefore, it is probably true, as said by Tiegs, that during anamorphosis “the twelfth leg-bearing segment, unlike the four that arise before it, fails to separate from the cerus-bearing segment.” The last segment of the body is a small anal segment (A, G, J, 14). On each side it bears a conical tubercle supporting a long sensory hair, the structure being known as a trichobothrium (trb). The anus (G, An) is situated apically between slightly protruding dorsal and ventral lobes.
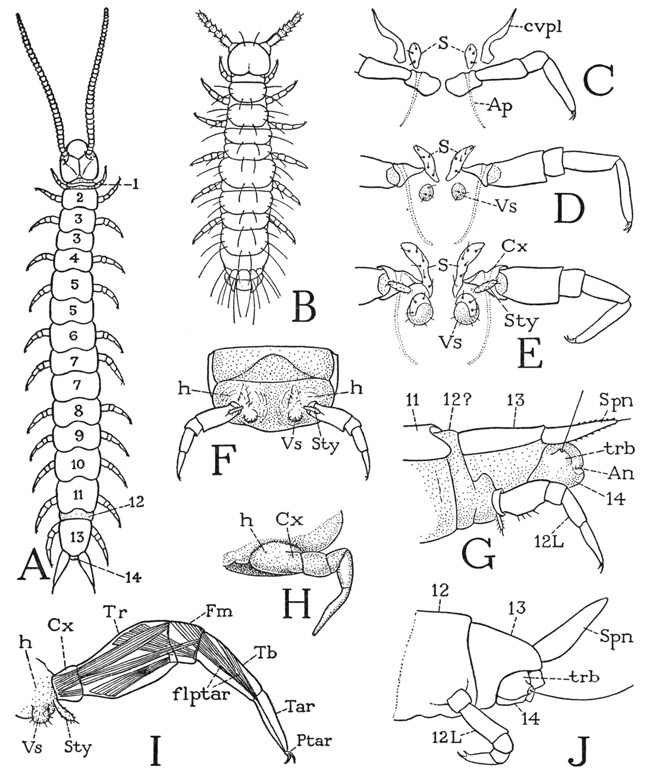
Fig. 71. Symphyla. General structure of the body and the legs.
A, Scutigerella immaculata (Newp.), adult. B, same, first instar (from Michelbacher, 1938). C, first legs and associated ventral sclerites. D, same, second legs, with imperfect vesicles. E, same, third legs, with styli and fully developed vesicles. F, same, ventral surface of segment from middle part of body. G, same, posterior end of body. H, Hanseniella agilis Tiegs, embryonic leg (from Tiegs, 1947). I, Scutigerella immaculata (Newp.), leg of adult, showing principal muscles and associated vesicle and stylus. J, Symphylella sp., posterior end of body with fully developed twelfth segment (from Tiegs, 1947).
For explanation of lettering see page 270.
It is a remarkable thing, shown by Tiegs (1940) in his study of the development of Hanseniella, that the anal segment in the embryo contains both coelomic sacs and nerve ganglia. The end segment of the symphylan, therefore, is not the telson, as it might appear to be; the telson, if present, must then be represented only by the anal lobes. The trichobothria, Tiegs says, arise as blunt outgrowths on the sides of the anal segment and appear to be vestiges of the appendages of this segment. The Symphyla evidently have 14 body segments representing primary somites, with segments 12 and 13 usually united.
The postembryonic development of the symphylans is anamorphic, the first instar having only nine body segments and either six (fig. 71 B) or seven pairs of legs. The preanal segment of the first instar is relatively large, and it is within this segment, as shown both by Michelbacher (1938) and by Tiegs (1940), that the new segments will be formed. From the coelomic sacs of the preanal segment, Tiegs says, arises the mesoderm for the segments that appear, during anamorphosis, between the seventh segment and the definitive preanal segment. The anamorphic development of the Symphyla thus differs from the teloblastic development of segments in other anamorphic arthropods, in which the generative zone lies immediately in front of the telson. It would appear, therefore, that in the symphylans telogenesis must already have produced before hatching the complete material for the segments added during postembryonic growth, and that anamorphosis in these arthropods is merely a delayed differentiation of this material into segments, accompanied by the formation of the new appendages. If the postembryonic development in Symphyla is to be explained in this manner, it evidently represents an intermediate stage between anamorphosis with typical teloblastic growth and epigenesis, or the completion of segmentation before hatching.
The Head and the Mouth Parts
The symphylan head (fig. 72 A, B, E, G) is of the prognathous type of structure insofar as the mouth parts lie horizontally against its undersurface and are directed forward, in which respect it resembles the head of a chilopod, diplopod, or pauropod. The head of Scutigerella is somewhat flattened (B); that of Hanseniella (E) is thicker and strongly declivous anteriorly. On its dorsal surface the head carries the long, many-segmented antennae, which are set in large membranous areas containing also the postantennal organs of Tömösvary (B, E, OT). Each antenna is pivoted on an articular process, or antennifer (A, af), arising from the cranial wall mesad of its base. Anterior to the antennae is a large epistomal area of the cranium (Epst), but there is no specifically demarked clypeus. The bilobed, spiny labrum (C, Lm) is scarcely separated from the epistome. The subantennal lateral margin of the cranium is formed by a narrow sclerotic bar (B, E) extending from the angle of the epistome to the posterior part of the cranium. In Scutigerella (B) the bar becomes so weak that its posterior cranial connection can be seen only in a well-stained specimen; in Hanseniella (E) the bar is somewhat thicker. Below this subantennal bar the cranium is deeply emarginate, as in the diplopods, but in the symphylan a wide membranous space intervenes between the bar and the base of the mandible (B, E) in which is one of the pair of head spiracles (Sp), the only tracheal apertures of the Symphyla. The dorsum of the head of Scutigerella immaculata (A) is marked by a midcranial sulcus that forks anteriorly to the antennal pivots and forms internally a corresponding Y-shaped ridge. From near the posterior end of the midcranial sulcus two pale lines diverge forward to the antennal membranes behind the organs of Tömösvary, but these lines are not grooves, nor do they form internal ridges. In stained specimens they are seen as faint, clear tracts of the cuticle, apparently of denser texture than the rest, devoid of the small setae that cover the other parts of the head; they are not present in other genera. The lines on the head of Scutigerella have nothing to do with moulting. The Symphyla have no eyes.
The antennae of Symphyla have been shown by Imms (1939) to be truly segmented appendages, since each division of the shaft is independently musculated from the one proximal to it. The basal segment contains only a single muscle; each of the others except the apical segment contains four muscles inserted on the segment beyond. According to Michelbacher (1938), the antenna of the first instar of Scutigerella immaculata (fig. 71 B) has a basal stalk and four free distal segments, but within the stalk are seven fully formed segments merely enclosed in a sheath. With subsequent moults, and the animal continues to moult throughout life, new segments are added. The average number of antennal segments in the adult is perhaps 30 or 40, but Michelbacher reports an observed maximum number of 60. When part of an antenna, or even the whole appendage, is broken off, the lost part is rapidly regenerated at the moults. From Michelbacher’s account of the antennal growth in Scutigerella it would seem that the new or regenerated segments are formed in the basal part of the appendage. Imms (1940), on the other hand, has shown that growth of the segmented antenna of the collembolan Campodea is acrogenous, that is, the new segments are cut off from the base of the permanent apical segment. This type of segment multiplication, however, would not allow of regeneration in an antenna lacking the distal part, unless each segment is capable of assuming the role of an apical segment when occasion demands.
The postantennal Tömösvary organs of Scutigerella (fig. 72 A, B, E, OT) have been well described and illustrated by Pflugfelder (1933). Each organ has a dome-shaped outer surface with a hole in the center that opens into a large globular chamber lined with a thin cuticle. From the floor of the chamber project numerous fine filaments that contain nerve fibrils given off from the outer ends of a group of sublying sensory nerve cells. In Hanseniella, as described by Tiegs (1940), the outer dome of the organ has a wide central opening, and the floor of the cavity beneath it is covered by a thick layer of gelatinous substance. The filaments are grouped in a central bundle that projects through the gelatinous layer; they are said to be cuticular tubules that resist boiling in caustic potash and are shed at each moult. Most writers find that the centripetal nerves of the Tömösvary organs go to the protocerebrum. Many sensory functions have been theoretically attributed to the organs of Tömösvary, but none has been experimentally demonstrated.
The head spiracles give rise to a tracheal system which is most elaborately developed in the head and extends back through only the first three segments of the body. The standard, widely copied illustration of the tracheal system of Scutigerella immaculata (see Verhoeff, 1933, Michelbacher, 1938) is taken from Haase (1884). According to Tiegs (1940), the tracheal invaginations of Hanseniella appear during the ninth day of embryonic development, apparently on the mandibular segment, but the openings become closed and remain so until the first larval moult. Presumably a complete tracheation of the body is not necessary on account of the small size of the animal and the permeability of its soft integument. The intima of the tracheal tubes of Scutigerella is said by Richards and Korda (1950) to have reticulate thickenings sometimes taking the form of incomplete taenidia.
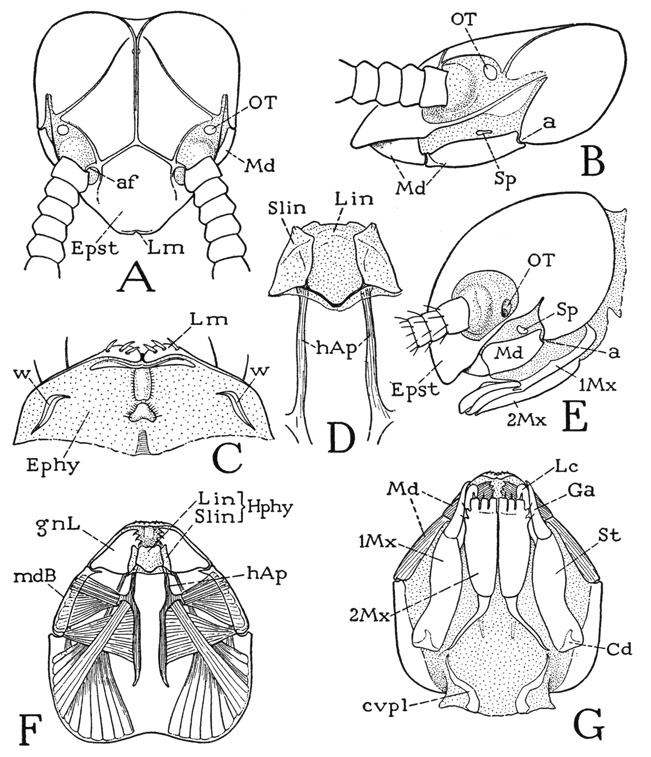
Fig. 72. Symphyla. The head.
A, Scutigerella immaculata (Newp.), head, dorsal. B, same, head, lateral. C, same, epipharyngeal surface and labrum, ventral. D, same, hypopharynx and head apodemes, dorsal. E, Hanseniella agilis Tiegs, head, lateral (outline from Tiegs, 1940). F, Scutigerella immaculata (Newp.), horizontal optical section of head below level of mandibles, ventral, showing mandibular muscles. G, same, head, ventral.
For explanation of lettering see page 270.
The ventrolateral parts of the symphylan head are occupied by the bases of the mandibles (fig. 72 B, E, Md), which latter thus have the same position as in the diplopods (fig. 63 D), though they do not occupy the entire lateral emarginations of the cranium. The ventral wall of the head (fig. 72 G) is membranous except as it is occupied by the two pairs of maxillae (1Mx, 2Mx), the second pair of which is partly united to form a labium. The mouth lies some distance behind the labrum, so that there is a long intervening epipharyngeal area (C, Ephy), as in the diplopods (fig. 63 E) and in Scutigera among the chilopods (fig. 54 D). Immediately behind the labrum of Scutigerella (fig. 72 C) is a transverse epipharyngeal ridge, followed by a median elevation. Laterally on each side is a strongly sclerotic, sinuous sclerite (w), which has a mechanical relation to a small process on the base of the gnathal lobe of the corresponding mandible (fig. 73 A, c). The two epipharyngeal sclerites of Scutigerella, therefore, are suggestive of the epipharyngeal arms of the fultural sclerites of the chilopods (fig. 54 D, w) on which the mandibles have loose articulations.
Projecting from beneath the mouth is a three-lobed hypopharynx (fig. 72 D; F, Hphy). Borrowing terms from entomology, the median hypopharyngeal lobe is the lingua (Lin), the lateral lobes the superlinguae (Slin), but the superlinguae evidently represent the paragnaths of Crustacea. In Scutigerella (D) the broad lingua is overlapped dorsally on its sides by the superlinguae, and the bases of the latter are connected by an angulated bar through the base of the lingua. According to Tiegs (1940), the superlinguae of Hanseniella develop on the sternum of the mandibular segment and are innervated from the mandibular ganglion. The lingua (“hypopharynx”) when first formed is purely a product of the first maxillary segment, but later “the labial ectoderm seems to become incorporated into its inferior wall.” In the Symphyla there are no premandibular sternal sclerites (fulturae) supporting the hypopharynx, but at the sides of the lingua arises a pair of strong head apodemes (D, F, hAp) that extend far back in the head cavity and give attachment to adductor muscles of the mouth parts and to ventral muscles from the body. The head apodemes of Symphyla are clearly homologues of the similar apodemes in Chilopoda, Diplopoda, and Pauropoda, though supporting sclerites are absent. The apodemes (hypopharyngeal apophyses) Tiegs says are developed in Hanseniella a little median to the bases of the mandibles, and later take a more median position; Tiegs assigns them to the mandibular segment. Hansen (1930, p. 270) mistook the head apodemes of Symphyla for ventral surface sclerites of the head, such as those present in the apterygote hexapods Diplura and Collembola.
The symphylan mouth parts (fig. 73) may be likened to those of insects in that the first maxillae (B) have something of the insect type of structure, and the second maxillae (C) are partly united to form a labiumlike underlip attached to the head. The mandibles (A), however, closely resemble the mandibles of a diplopod, since each consists of a basal plate implanted on the side of the head and of an independently movable gnathal lobe.
The mandibles of Symphyla are relatively shorter than those of the diplopods, and, as noted above, their basal plates do not occupy the entire lateral emarginations of the cranium (fig. 72, B, E). The gnathal lobes (fig. 73 A, gnL) are flattened and lie horizontally in the preoral cavity of the head; each is articulated to the basal plate (mdB) by the outer angle of its base and has a process (c) on its dorsal surface that loosely bears on the corresponding lateral sclerite of the epipharynx (fig. 72 C, w). The mandibular musculature includes the same muscles that are attached on the diplopod jaw, but the two ventral adductors (fig. 73 A, 4a, 4b) both arise on the head apodeme (hAp), there being no intergnathal ligament such as that of the diplopods. A dorsal muscle (2) from the posterior part of the cranium is attached on the mandibular base; the gnathal lobe has a huge cranial flexor (1). Though the movable gnathal lobe of the symphylan mandible, with its cranial flexor muscle, resembles the movable lacinea of a generalized insect maxilla, it is hardly to be supposed that the symphylan or diplopod mandible represents a primitive arthropod jaw. The “segmentation” of the mandible of Hanseniella, according to Tiegs, takes place on the ninth day of embryonic growth.
The maxillae of Scutigerella are relatively long appendages lying laterally against the underside of the head (fig. 72 G, 1Mx). Each has an elongate, elaborately musculated basal plate, or stipes (fig. 73 B, St), attached by its entire length to the head wall, and two free apical lobes which may be termed galea (Ga) and lacinia (Lc), since they appear to correspond with the lobes so named in an insect maxilla. The lacinia has a cranial flexor muscle. A small lateral spur (Plp) on the stipes near the base of the galea is possibly a remnant of the palpus. The implantation of the basal plates of the mandibles and maxillae on the head wall, Tiegs (1940) observes, “does not appear seriously to impede their movement,” for “if an animal is watched under a lens when feeding, the ventro-lateral wall of the head is seen in active movement.” The strong musculature of the basal plates of the two appendages would imply that this movement is of considerable importance.
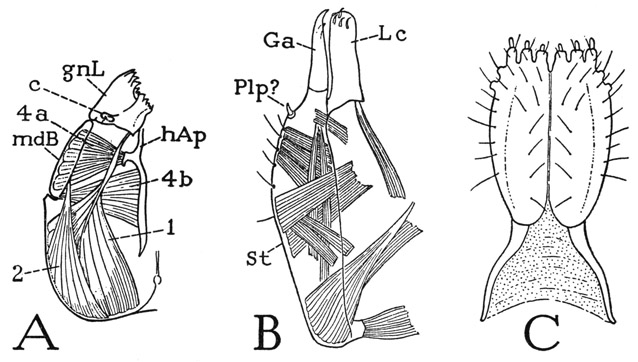
Fig. 73. Symphyla. The mouth parts of Scutigerella immaculata (Newp.).
A, left mandible and muscles, dorsal. B, left first maxilla, dorsal. C, second maxillae, labium, ventral.
For explanation of lettering see page 270.
The labium of Symphyla (fig. 73 C) is evidently composed of two head appendages homologous with the second maxillae of the chilopods or the labial components of insects, though it has no resemblance in form to either. It consists of a broad, oval anterior plate, divided by a median groove, with six papilla-bearing lobes on its distal margin, and of two slender, tapering proximal arms diverging posteriorly in the membranous head wall to the anterior ends of a pair of cervical sclerites (fig. 72 G, cvpl). Palpi are entirely absent. In the embryonic development of Hanseniella, Tiegs (1940) says, “the most anterior intersegmental cleft is between the maxillary and labial segments,” the labial segment being at first clearly a part of the body and only in late embryonic growth becoming a part of the head. Considering the lack of evidence of a second maxillary segment in the head of the diplopods and pauropods, it seems probable that the common ancestors of the progoneate myriapods had only one maxillary segment incorporated into the head.
The Body Appendages
The legs of Symphyla, except those of the first pair, are all alike, and each is divided into six segments (fig. 71 I). The segments are generally interpreted as being a small coxa (Cx), an unusually large trochanter (Tr), a very short femur (Fm), an elongate tibia (Tb), an undivided tarsus (Tar), and a clawlike pretarsus (Ptar) with an accessory lateral claw. The four distal segments are fairly well identified by the distribution of the fibers of the flexor muscle of the pretarsus (flptar), but the identity of the two proximal segments may be questioned. The large second segment, however, is suggestive of the long second trochanter of the diplopod leg (fig. 65 H). If the basal segment of the symphylan leg is the coxa, as it appears to be, then a first trochanter is absent. The symphylid leg has some resemblance to the leg of a pauropod (fig. 70 I) but none at all to an insect leg (fig. 83 A), in which the principal segment proximal to the “knee” is the femur and the trochanter is always small. As before noted, the relative size of the leg segments among the arthropods generally is no index of the identity of the segments. The legs of the first pair in the Symphyla (fig. 71 C) are not only smaller than the others, but they lack a segment which apparently is the tibia. In some species these legs are much reduced in size; in others they are almost as long as the other legs.
Each leg of the Symphyla arises from a slightly bulging area of the lateroventral wall of the supporting body segment (fig. 71 F, I, h), which bears also on most of the segments a short stylus (Sty) and an eversible vesicle (Vs). The leg-bearing mound has been interpreted by Hansen (1930) as the coxa of the leg and by Ewing (1928) as a subcoxa. Tiegs (1940) calls it the “limb-base,” and in his figure of an embryonic leg of Hanseniella (H) the “limb-base” (h) does have the appearance of a basal segment of the limb, but Tiegs asserts that it does not develop as a part of the leg and is therefore not a subcoxal segment of the leg. It represents the area of a “ventral organ,” that is, one of the paired segmental thickenings of the embryonic ventral ectoderm from the inner surface of which the nerve tissue is differentiated. The eversible vesicle, according to Tiegs, is the epidermal remnant of the ventral organ protruded at the surface. The stylus arises laterad of the vesicle very close to the base of the coxa, but not on it. Vesicles and styli are absent on the first body segment (C); on the second segment (D) is a pair of imperfect vesicles; on the third (E) and the following segments are both styli and fully developed vesicles. The vesicles are eversible and are said to have retractor muscles, but the styli lack muscles. The function of these organs is not known. The coxae are articulated ventrally on small sternal plates lying before the vesicles (D, E, S), but have no dorsal articulations.
The spinnerets of the preanal segment (fig. 71 G, J, Spn) give exit at their tips to a pair of glands, the secretion of which hardens on contact with the air to a silklike thread. Because of their prominence at the end of the body (A) the spinnerets have been thought to be homologues of the insect cerci. The preanal segment of the Symphyla is at most the thirteenth body segment; while it is true that the cerci of many adult insects are attached on the thirteenth postcephalic segment, embryologists are agreed that they are the appendages of the fourteenth segment in the embryo. Hence, though the insect cerci may also contain glands, as in some of the Japygidae, their segmental status is not the same as that of the symphylan spinnerets. Among the diplopods, members of the Nematophora have silk glands that open through spines on the anal segment, but in the arthropods generally silk-spinning is a function of such widely different appendages that it cannot be invoked as evidence of homology in any particular case. If the trichobothria of the anal segment of the symphylans are the appendages of this, the fourteenth, body segment, they, rather than the spinnerets, would be the homologues of the insect cerci. Of just what use the silk spun from the spinnerets is to the symphylans in nature is not known, but Michelbacher (1938) says that, when ejected on a camel-hair brush, the threads form a most convenient means of transferring live specimens without injury from one container to another.
Life History of Scutigerella immaculata
The life history of the garden centipede is probably most fully and correctly described by Michelbacher (1938). The first instar (fig. 71 B) has six pairs of legs; Tiegs (1940) says there are seven pairs of legs in the first instar of Hanseniella. According to Michelbacher, Scutigerella immaculata goes through seven immature instars, adding a pair of legs at each moult, the adult number of 12 being thus attained by the seventh instar. The definitive number of back plates, however, is present in the sixth instar. During the immature stages the antennae have increased in length by the successive addition of segments until in the young adult there are about 30 antennal segments. The symphylids, however, do not cease moulting on becoming adult. Michelbacher says that evidence indicates the number of moults may be in excess of 50. During the first few adult moults the body may increase in size up to a maximum length of 7 or 8 mm., but the antennae continue to add segments throughout life until each may have as many as 60 segments. Under laboratory conditions, Michelbacher says, the garden centipede lives for several years.
The manner of ecdysis in the Symphyla is the same as that in other myriapods; there is no lengthwise dorsal rupture of the cuticle forking on the head, as is usual with insects. The outer cuticle splits transversely behind the head and the animal crawls out of the body skin. Williams (1907) says this is at least the most common method of ecdysis, and the one observed in a specimen in the act of emergence, but that shed skins are often found to be torn in various ways. Michelbacher (1938) also observes that at ecdysis “the garden centipede frees its head from the capsule and by forward propulsions starts to work its way out of the skin.”
Explanation of Lettering on Figures 71-73
a, cranial articulation of mandible. af, antennifer.
An, anus.
Ap, apodeme.
c, epipharyngeal articulation of mandible.
Cd, cardo.
cvpl, cervical plate.
Cx, coxa.
Ephy, epipharyngeal surface.
Epst, epistome.
flptar, flexor muscle of pretarsus.
Fm, femur.
Ga, galea
gnL, gnathal lobe of mandible.
h, lobe of body supporting the leg.
hAp, head apodeme, hypopharyngeal apophysis.
Hphy, hypopharynx.
L, leg.
Lc, lacinia.
Lin, lingua.
Lm, labrum.
Md, mandible.
mdB, base of mandible.
1Mx, first maxilla.
2Mx, second maxilla.
OT, organ of Tömösvary.
Plp?, small spur of maxilla, possibly vestige of palpus.
Ptar, pretarsus.
S, sternum
Slin, superlingua.
Sp, spiracle.
Spn, spinneret.
St, stipes.
Sty, stylus.
Tar, tarsus.
Tb, tibia.
Tr, trochanter.
trb, trichobothrium.
Vs, eversible vesicle.
w, lateral epipharyngeal sclerite.